

Active FOXO1 Is a Key Determinant of Isoform-Specific Progesterone Receptor Transactivation and Senescence Programming
- PMID: 26577046
- PMCID: PMC4755896
- DOI: 10.1158/1541-7786.MCR-15-0431
Active FOXO1 Is a Key Determinant of Isoform-Specific Progesterone Receptor Transactivation and Senescence Programming
Abstract
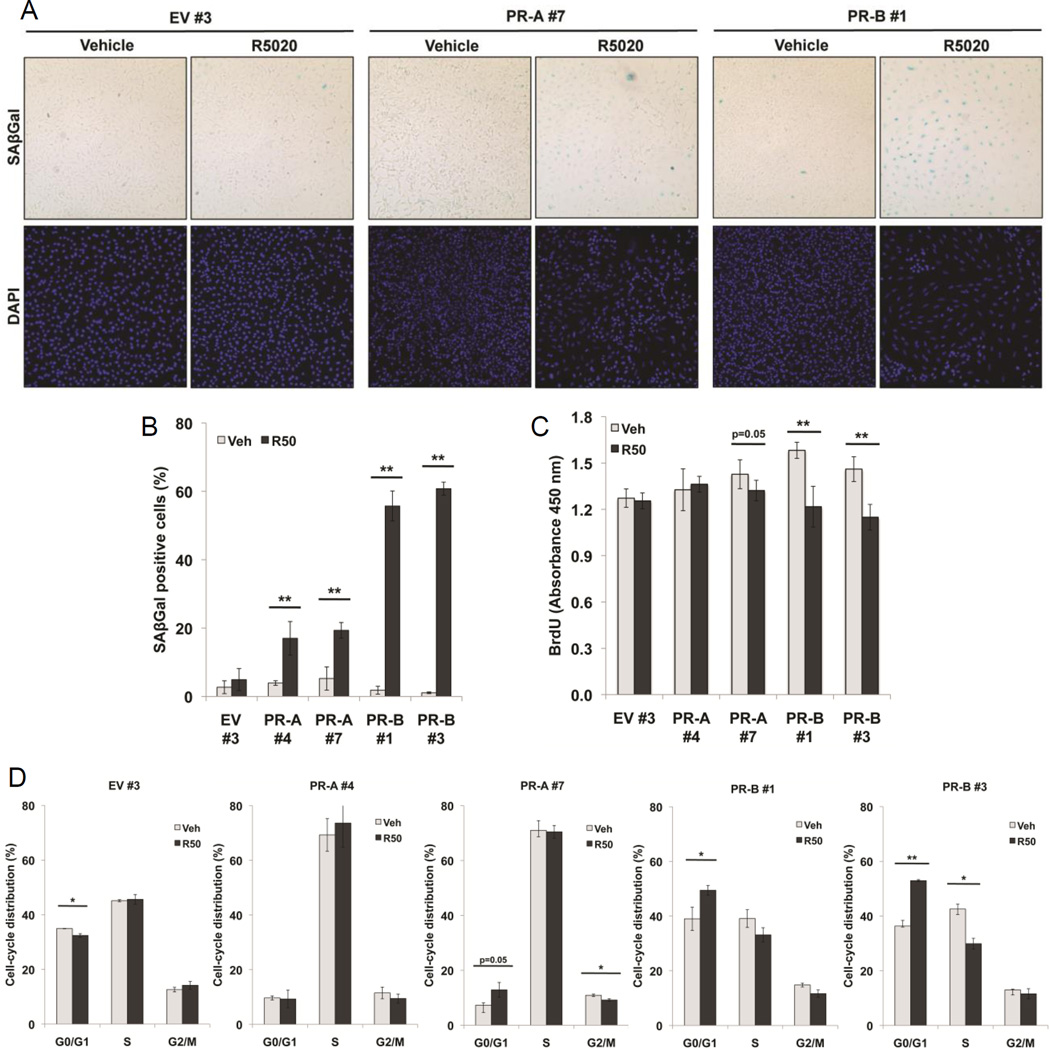
Progesterone promotes differentiation coupled to proliferation and prosurvival in the breast, but inhibits estrogen-driven growth in the reproductive tract and ovaries. Herein, it is demonstrated, using progesterone receptor (PR) isoform-specific ovarian cancer model systems, that PR-A and PR-B promote distinct gene expression profiles that differ from PR-driven genes in breast cancer cells. In ovarian cancer models, PR-A primarily regulates genes independently of progestin, while PR-B is the dominant ligand-dependent isoform. Notably, FOXO1 and the PR/FOXO1 target gene p21 (CDKN1A) are repressed by PR-A, but induced by PR-B. In the presence of progestin, PR-B, but not PR-A, robustly induced cellular senescence via FOXO1-dependent induction of p21 and p15 (CDKN2B). Chromatin immunoprecipitation (ChIP) assays performed on PR isoform-specific cells demonstrated that while each isoform is recruited to the same PRE-containing region of the p21 promoter in response to progestin, only PR-B elicits active chromatin marks. Overexpression of constitutively active FOXO1 in PR-A-expressing cells conferred robust ligand-dependent upregulation of the PR-B target genes GZMA, IGFBP1, and p21, and induced cellular senescence. In the presence of endogenous active FOXO1, PR-A was phosphorylated on Ser294 and transactivated PR-B at PR-B target genes; these events were blocked by the FOXO1 inhibitor (AS1842856). PR isoform-specific regulation of the FOXO1/p21 axis recapitulated in human primary ovarian tumor explants treated with progestin; loss of progestin sensitivity correlated with high AKT activity.
Implications: This study indicates FOXO1 as a critical component for progesterone signaling to promote cellular senescence and reveals a novel mechanism for transcription factor control of hormone sensitivity.
©2015 American Association for Cancer Research.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures








Similar articles
-
Progesterone receptors induce FOXO1-dependent senescence in ovarian cancer cells.Cell Cycle. 2013 May 1;12(9):1433-49. doi: 10.4161/cc.24550. Epub 2013 Apr 10. Cell Cycle. 2013. PMID: 23574718 Free PMC article.
-
Progesterone receptor-B regulation of insulin-like growth factor-stimulated cell migration in breast cancer cells via insulin receptor substrate-2.Mol Cancer Res. 2008 Sep;6(9):1491-8. doi: 10.1158/1541-7786.MCR-07-2173. Mol Cancer Res. 2008. PMID: 18819936 Free PMC article.
-
Linkage of progestin and epidermal growth factor signaling: phosphorylation of progesterone receptors mediates transcriptional hypersensitivity and increased ligand-independent breast cancer cell growth.Steroids. 2007 Feb;72(2):188-201. doi: 10.1016/j.steroids.2006.11.009. Epub 2006 Dec 14. Steroids. 2007. PMID: 17173941 Free PMC article.
-
The role and mechanism of progesterone receptor activation of extra-nuclear signaling pathways in regulating gene transcription and cell cycle progression.Steroids. 2008 Oct;73(9-10):922-8. doi: 10.1016/j.steroids.2008.01.010. Epub 2008 Jan 19. Steroids. 2008. PMID: 18321550 Review.
-
Progesterone receptor action: translating studies in breast cancer models to clinical insights.Adv Exp Med Biol. 2008;630:94-111. Adv Exp Med Biol. 2008. PMID: 18637487 Review.
Cited by
-
Progesterone receptor isoforms, agonists and antagonists differentially reprogram estrogen signaling.Oncotarget. 2017 Sep 28;9(4):4282-4300. doi: 10.18632/oncotarget.21378. eCollection 2018 Jan 12. Oncotarget. 2017. PMID: 29435103 Free PMC article.
-
Phosphorylated Progesterone Receptor Isoforms Mediate Opposing Stem Cell and Proliferative Breast Cancer Cell Fates.Endocrinology. 2019 Feb 1;160(2):430-446. doi: 10.1210/en.2018-00990. Endocrinology. 2019. PMID: 30597041 Free PMC article.
-
90 YEARS OF PROGESTERONE: Steroid receptors as MAPK signaling sensors in breast cancer: let the fates decide.J Mol Endocrinol. 2020 Jul;65(1):T35-T48. doi: 10.1530/JME-19-0274. J Mol Endocrinol. 2020. PMID: 32209723 Free PMC article. Review.
-
Utilizing an Endogenous Progesterone Receptor Reporter Gene for Drug Screening and Mechanistic Study in Endometrial Cancer.Cancers (Basel). 2022 Oct 6;14(19):4883. doi: 10.3390/cancers14194883. Cancers (Basel). 2022. PMID: 36230806 Free PMC article.
-
Dopamine-induced interactions of female mouse hypothalamic proteins with progestin receptor-A in the absence of hormone.J Neuroendocrinol. 2020 Oct;32(10):e12904. doi: 10.1111/jne.12904. Epub 2020 Sep 30. J Neuroendocrinol. 2020. PMID: 33000549 Free PMC article.
References
-
- Howlader N, Noone AM, Krapcho M, Garshell J, Miller D, Altekruse SF, Kosary CL, Yu M, Ruhl J, Tatalovich Z. SEER Cancer Statistics Review, 1975–2012. National Cancer Institute. 2015
-
- Edmondson RJ, Monaghan JM. The epidemiology of ovarian cancer. International journal of gynecological cancer : official journal of the International Gynecological Cancer Society. 2001;11:423–429. - PubMed
-
- Gabra H, Watson JE, Taylor KJ, Mackay J, Leonard RC, Steel CM, Porteous DJ, Smyth JF. Definition and refinement of a region of loss of heterozygosity at 11q23.3-q24.3 in epithelial ovarian cancer associated with poor prognosis. Cancer Res. 1996;56:950–954. - PubMed
-
- Kelsey JL, Gammon MD, John EM. Reproductive factors and breast cancer. Epidemiologic reviews. 1993;15:36–47. - PubMed
-
- Banks E, Beral V, Reeves G. The epidemiology of epithelial ovarian cancer: a review. International Journal of Gynecological Cancer. 1997;7:425–438.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
