

High yield extraction of pure spinal motor neurons, astrocytes and microglia from single embryo and adult mouse spinal cord
- PMID: 26577180
- PMCID: PMC4649473
- DOI: 10.1038/srep16763
High yield extraction of pure spinal motor neurons, astrocytes and microglia from single embryo and adult mouse spinal cord
Abstract
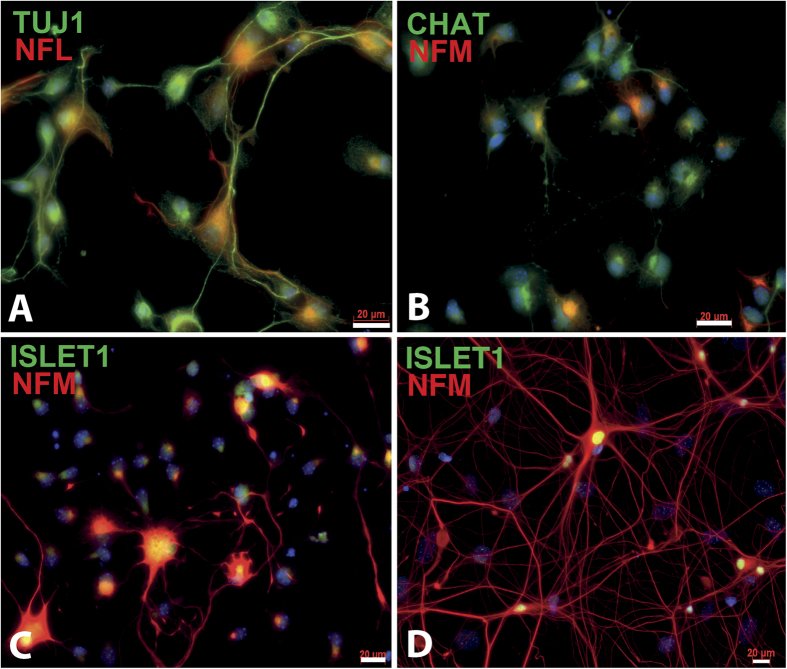
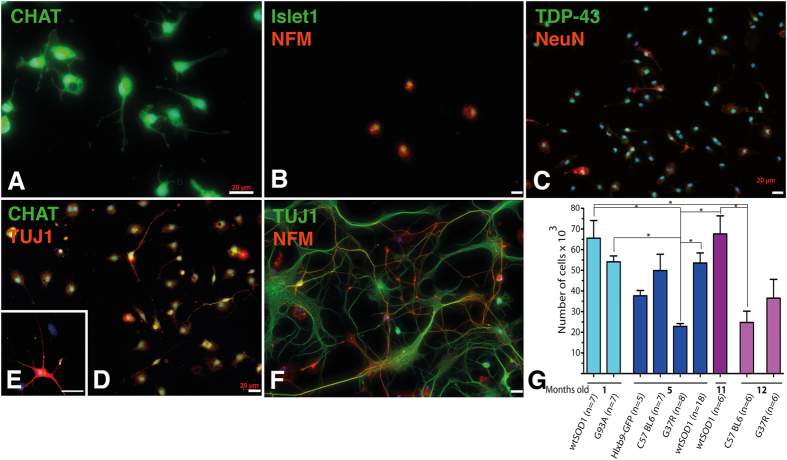
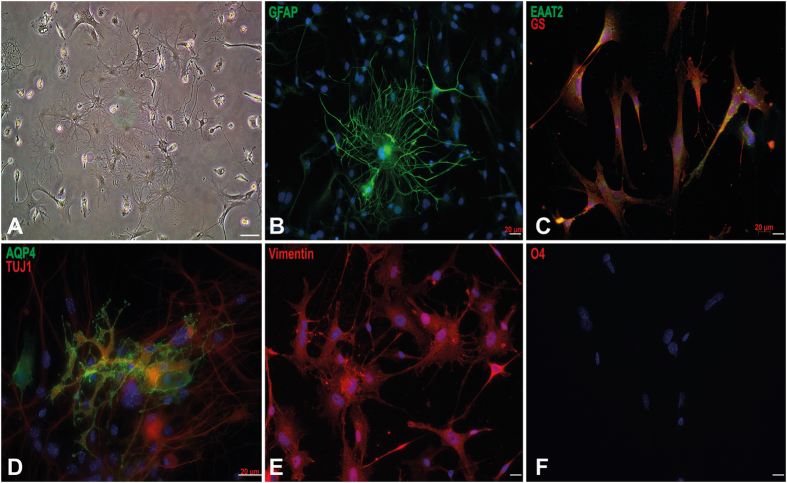
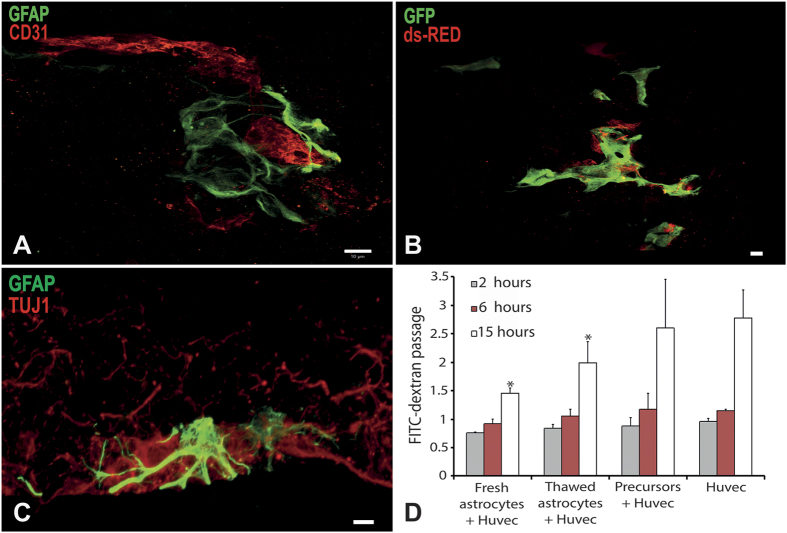
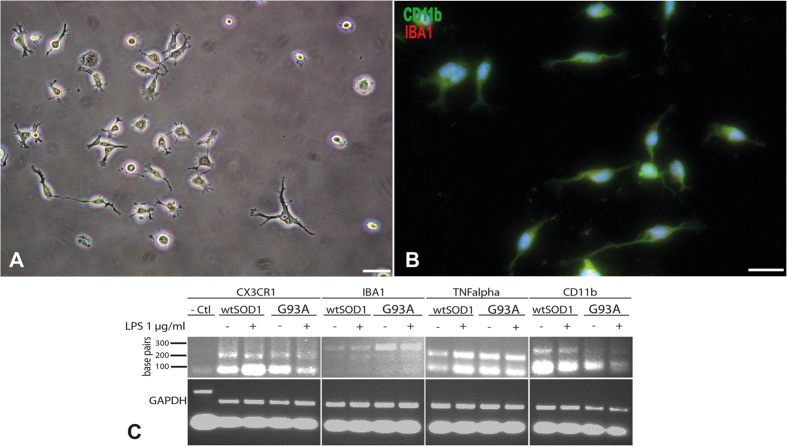
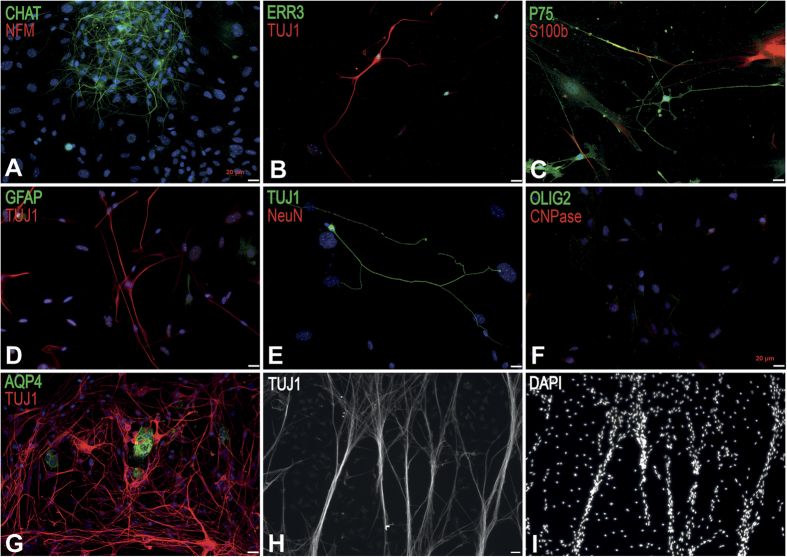
Extraction of mouse spinal motor neurons from transgenic mouse embryos recapitulating some aspects of neurodegenerative diseases like amyotrophic lateral sclerosis has met with limited success. Furthermore, extraction and long-term culture of adult mouse spinal motor neurons and glia remain also challenging. We present here a protocol designed to extract and purify high yields of motor neurons and glia from individual spinal cords collected on embryos and adult (5-month-old) normal or transgenic mice. This method is based on mild digestion of tissue followed by gradient density separation allowing to obtain two millions motor neurons over 92% pure from one E14.5 single embryo and more than 30,000 from an adult mouse. These cells can be cultured more than 14 days in vitro at a density of 100,000 cells/cm(2) to maintain optimal viability. Functional astrocytes and microglia and small gamma motor neurons can be purified at the same time. This protocol will be a powerful and reliable method to obtain motor neurons and glia to better understand mechanisms underlying spinal cord diseases.
Figures
References
-
- Bunton-Stasyshyn R. K., Saccon R. A., Fratta P. & Fisher E. M. SOD1 Function and Its Implications for Amyotrophic Lateral Sclerosis Pathology: New and Renascent Themes. Neuroscientist 21, 519–29 (2014). - PubMed
-
- Dion P. A., Daoud H. & Rouleau G. A. Genetics of motor neuron disorders: new insights into pathogenic mechanisms. Nature reviews. Genetics 10, 769–782 (2009). - PubMed
-
- Berthod F. & Gros-Louis F. in Amyotrophic Lateral Sclerosis. (ed. Maurer M. ) 81–124 (InTech, Rijeka, 2012).
-
- Gingras M., Gagnon V., Minotti S., Durham H. D. & Berthod F. Optimized protocols for isolation of primary motor neurons, astrocytes and microglia from embryonic mouse spinal cord. J. Neurosci. Methods 163, 111–118 (2007). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
