

CD4+ T effector memory cell dysfunction is associated with the accumulation of granulocytic myeloid-derived suppressor cells in glioblastoma patients
- PMID: 26578623
- PMCID: PMC4864257
- DOI: 10.1093/neuonc/nov280
CD4+ T effector memory cell dysfunction is associated with the accumulation of granulocytic myeloid-derived suppressor cells in glioblastoma patients
Abstract
Background: Myeloid-derived suppressor cells (MDSCs) comprise a heterogeneous population of myeloid cells that are significantly expanded in cancer patients and are associated with tumor progression.
Methods: Multicolor flow cytometry was used to study the frequency, phenotype, and function of MDSCs in peripheral blood and freshly resected tumors of 52 participants with primary glioblastoma (GBM).
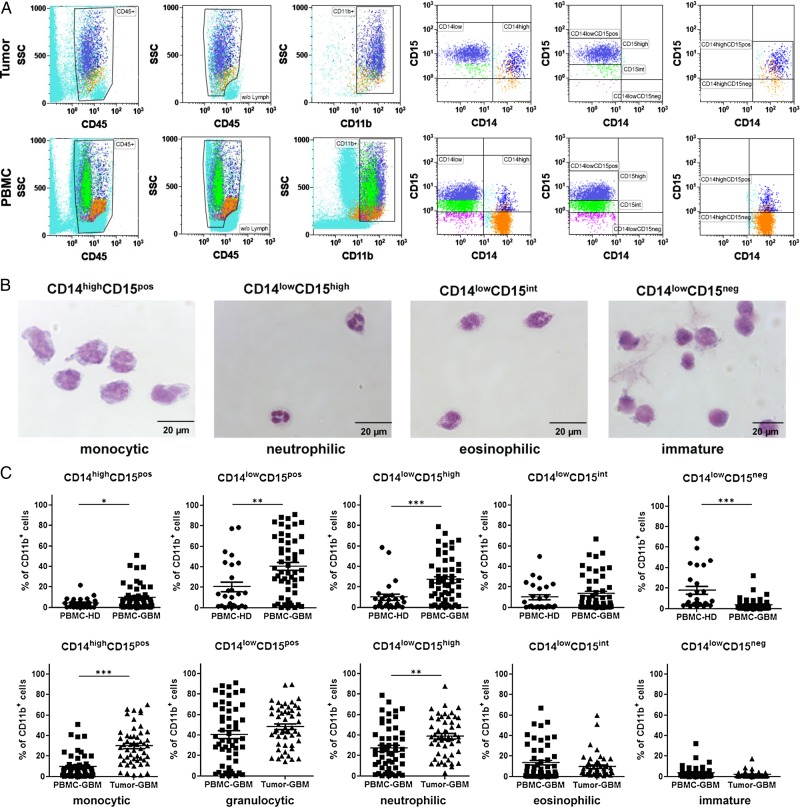
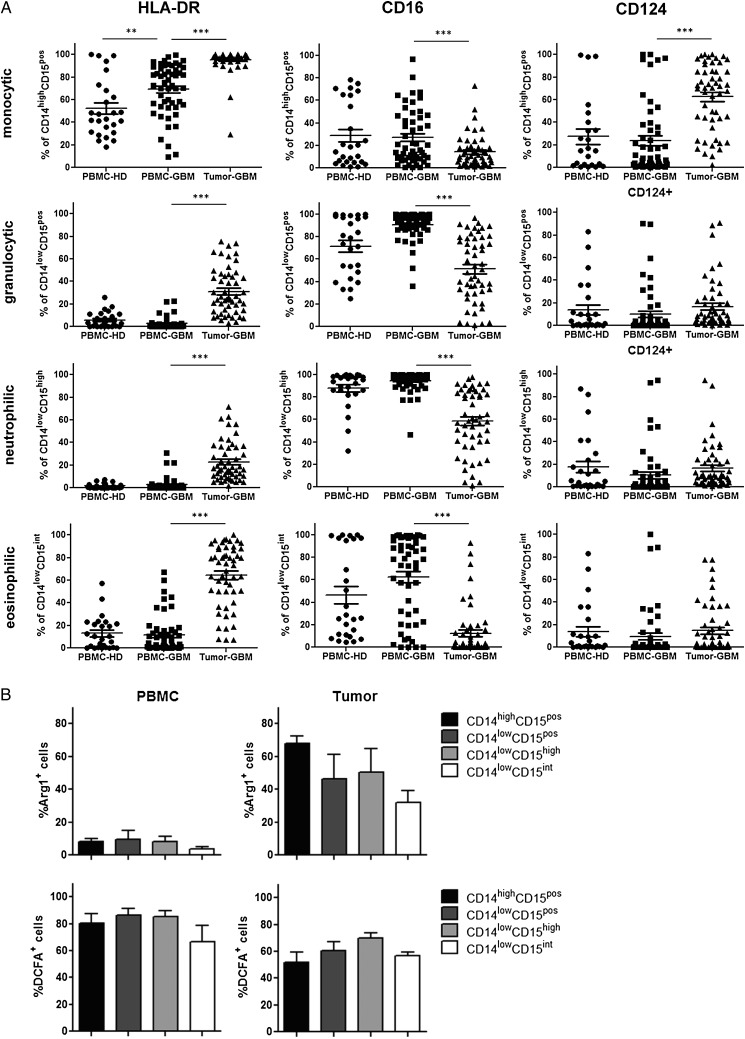
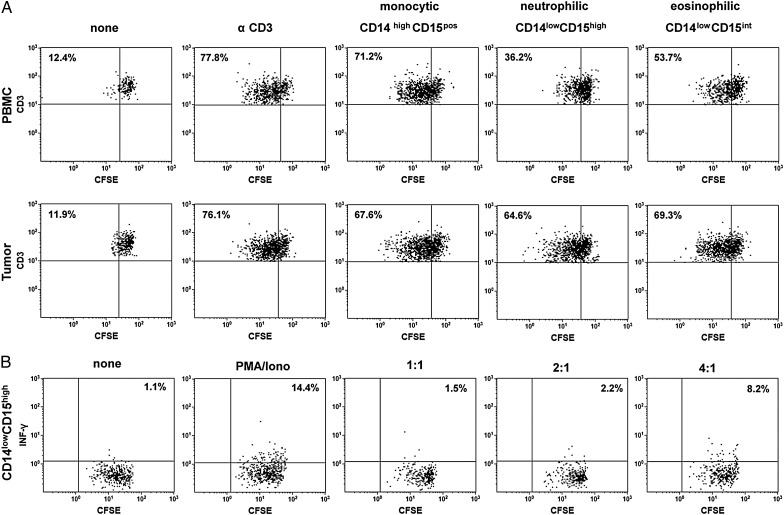
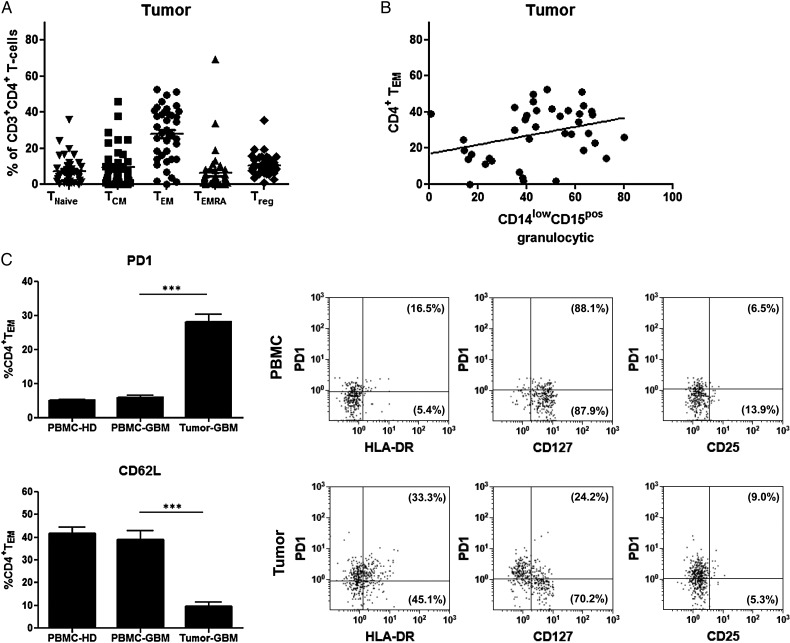
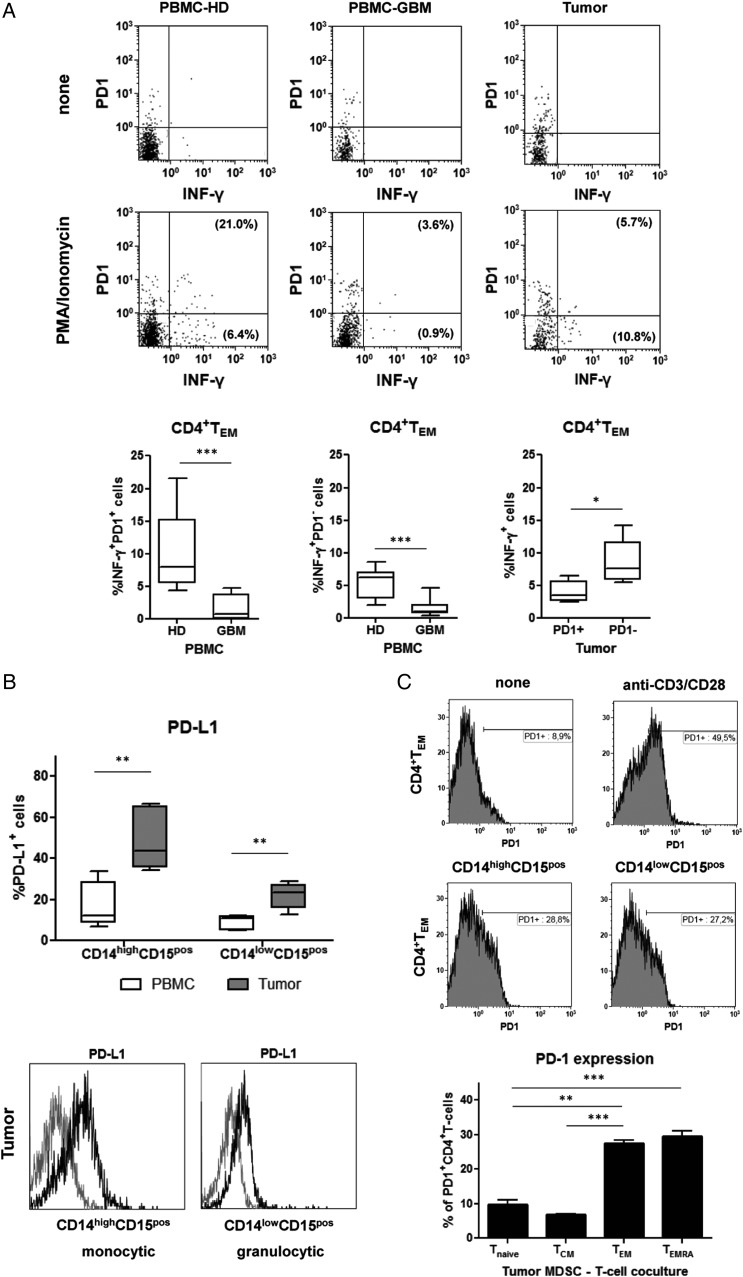
Results: The frequency of CD14(high)CD15(pos) monocytic and CD14(low)CD15(pos) granulocytic MDSCs was significantly higher in peripheral blood of GBM participants compared with healthy donors. The majority of granulocytic MDSCs consisted of CD14(low)CD15(high) neutrophilic MDSCs with high T-cell suppressive capacities. At the tumor side, we found an increase in CD14(high)CD15(pos) monocytic MDSCs and high frequencies of CD14(low)CD15(pos) granulocytic MDSCs that displayed an activated phenotype with downregulation of CD16 and upregulation of HLA-DR molecules, which did not inhibit T-cell proliferative responses in vitro. However, a strong association between granulocytic MDSCs and CD4(+) effector memory T-cells (TEM) within the tumors was detected. Tumor-derived CD4(+) TEM expressed high levels of PD-1 when compared with their blood-derived counterparts and were functionally exhausted. The respective ligand, PD-L1, was significantly upregulated on tumor-derived MDSCs, and T-cell co-culture experiments confirmed that glioma-infiltrating MDSCs can induce PD-1 expression on CD4(+) TEM.
Conclusions: Our findings provide a detailed characterization of different MDSC subsets in GBM patients and indicate that both granulocytic MDSCs in peripheral blood and at the tumor site play a major role in GBM-induced T-cell suppression.
Keywords: PD-1; PD-L1; T effector memory cell; glioblastoma; myeloid-derived suppressor cell.
© The Author(s) 2015. Published by Oxford University Press on behalf of the Society for Neuro-Oncology. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
Similar articles
-
Increase in both CD14-positive and CD15-positive myeloid-derived suppressor cell subpopulations in the blood of patients with glioma but predominance of CD15-positive myeloid-derived suppressor cells in glioma tissue.J Neuropathol Exp Neurol. 2015 May;74(5):390-400. doi: 10.1097/NEN.0000000000000183. J Neuropathol Exp Neurol. 2015. PMID: 25853692
-
Immunosuppressive myeloid-derived suppressor cells are increased in splenocytes from cancer patients.Cancer Immunol Immunother. 2017 Apr;66(4):503-513. doi: 10.1007/s00262-016-1953-z. Epub 2017 Jan 20. Cancer Immunol Immunother. 2017. PMID: 28108766 Free PMC article.
-
A circulating subpopulation of monocytic myeloid-derived suppressor cells as an independent prognostic/predictive factor in untreated non-small lung cancer patients.J Immunol Res. 2014;2014:659294. doi: 10.1155/2014/659294. Epub 2014 Nov 11. J Immunol Res. 2014. PMID: 25436215 Free PMC article.
-
On the origin of myeloid-derived suppressor cells.Oncotarget. 2017 Jan 10;8(2):3649-3665. doi: 10.18632/oncotarget.12278. Oncotarget. 2017. PMID: 27690299 Free PMC article. Review.
-
Explicating the Pivotal Pathogenic, Diagnostic, and Therapeutic Biomarker Potentials of Myeloid-Derived Suppressor Cells in Glioblastoma.Dis Markers. 2020 Nov 4;2020:8844313. doi: 10.1155/2020/8844313. eCollection 2020. Dis Markers. 2020. PMID: 33204365 Free PMC article. Review.
Cited by
-
Anti-PD-1 and Extended Half-life IL2 Synergize for Treatment of Murine Glioblastoma Independent of Host MHC Class I Expression.Cancer Immunol Res. 2023 Jun 2;11(6):763-776. doi: 10.1158/2326-6066.CIR-22-0570. Cancer Immunol Res. 2023. PMID: 36921098 Free PMC article.
-
Cancer-Derived VEGF-C Increases Chemokine Production in Lymphatic Endothelial Cells to Promote CXCR2-Dependent Cancer Invasion and MDSC Recruitment.Cancers (Basel). 2019 Aug 6;11(8):1120. doi: 10.3390/cancers11081120. Cancers (Basel). 2019. PMID: 31390756 Free PMC article.
-
Role of Neutrophils and Myeloid-Derived Suppressor Cells in Glioma Progression and Treatment Resistance.Int J Mol Sci. 2020 Mar 13;21(6):1954. doi: 10.3390/ijms21061954. Int J Mol Sci. 2020. PMID: 32182988 Free PMC article. Review.
-
Metabolic and functional reprogramming of myeloid-derived suppressor cells and their therapeutic control in glioblastoma.Cell Stress. 2019 Jan 23;3(2):47-65. doi: 10.15698/cst2019.02.176. Cell Stress. 2019. PMID: 31225500 Free PMC article. Review.
-
[The role of myeloid-derived suppressor cells in glioma microenvironment].Sheng Wu Yi Xue Gong Cheng Xue Za Zhi. 2019 Jun 25;36(3):515-520. doi: 10.7507/1001-5515.201806045. Sheng Wu Yi Xue Gong Cheng Xue Za Zhi. 2019. PMID: 31232558 Free PMC article. Review. Chinese.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
