

The structure of Medicago truncatula δ(1)-pyrroline-5-carboxylate reductase provides new insights into regulation of proline biosynthesis in plants
- PMID: 26579138
- PMCID: PMC4626632
- DOI: 10.3389/fpls.2015.00869
The structure of Medicago truncatula δ(1)-pyrroline-5-carboxylate reductase provides new insights into regulation of proline biosynthesis in plants
Abstract
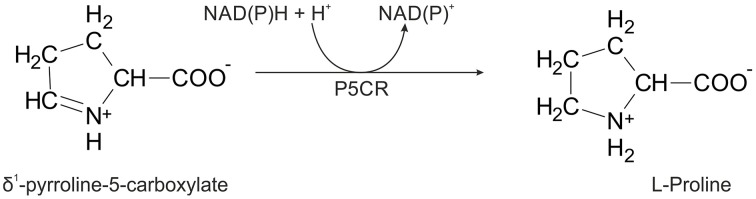
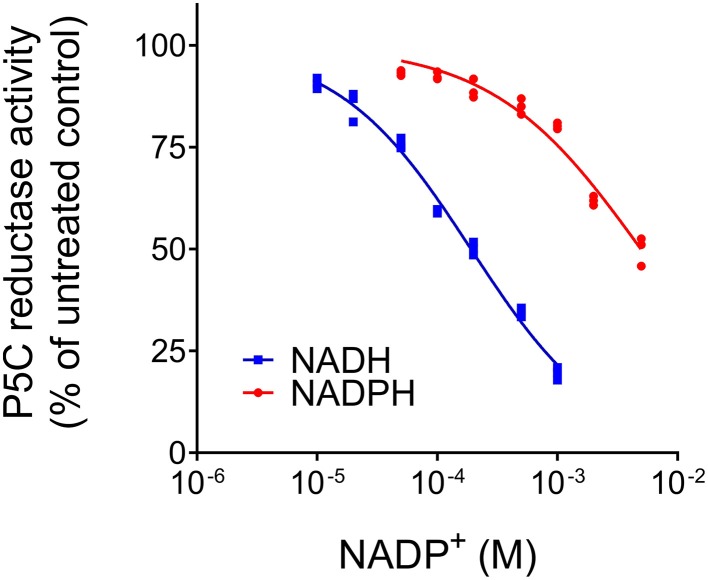
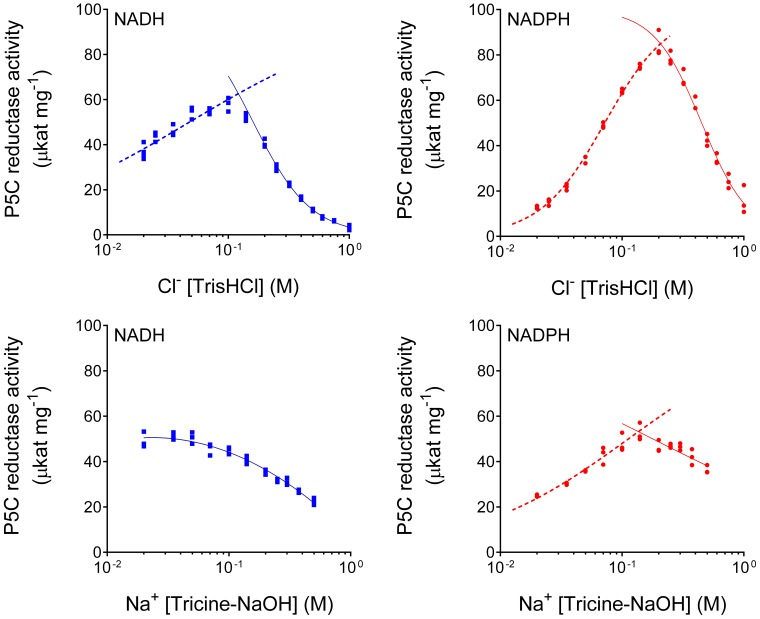
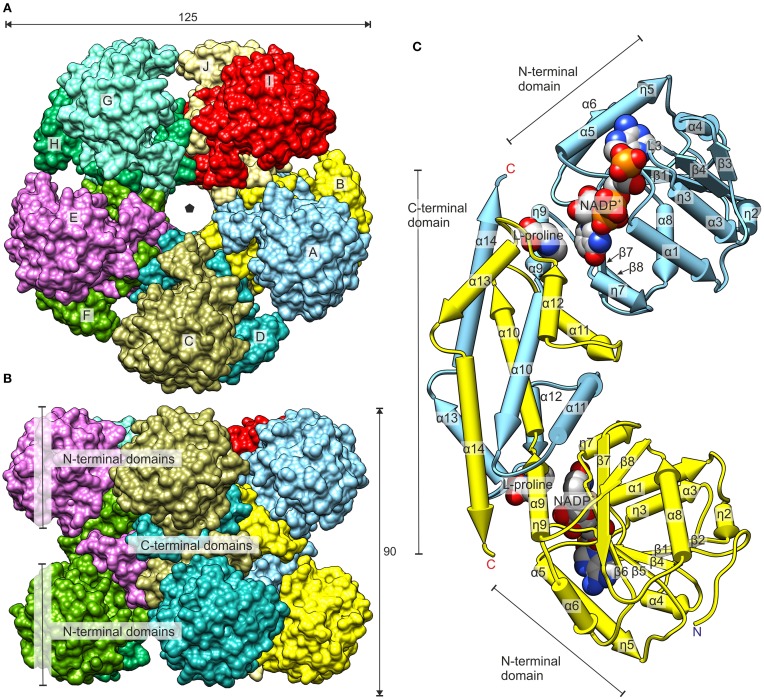
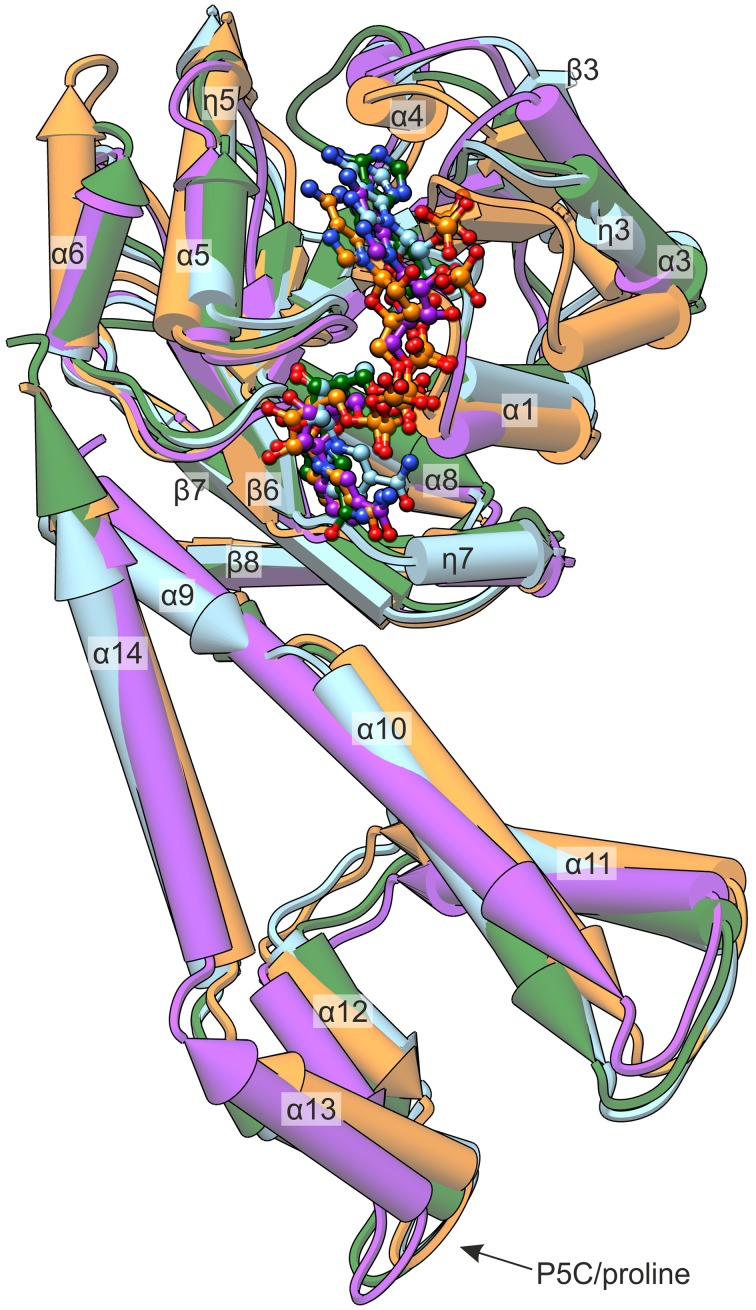
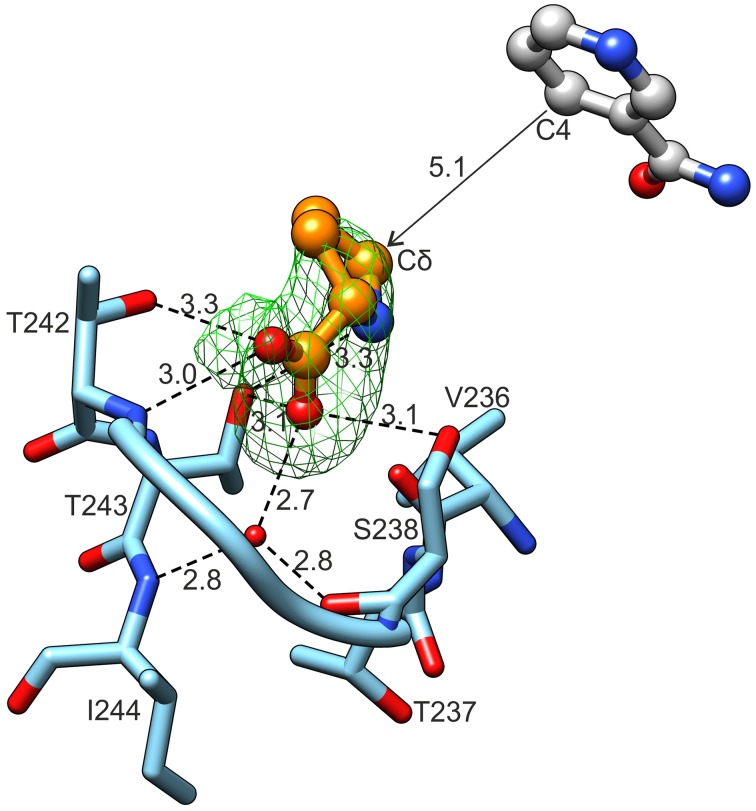
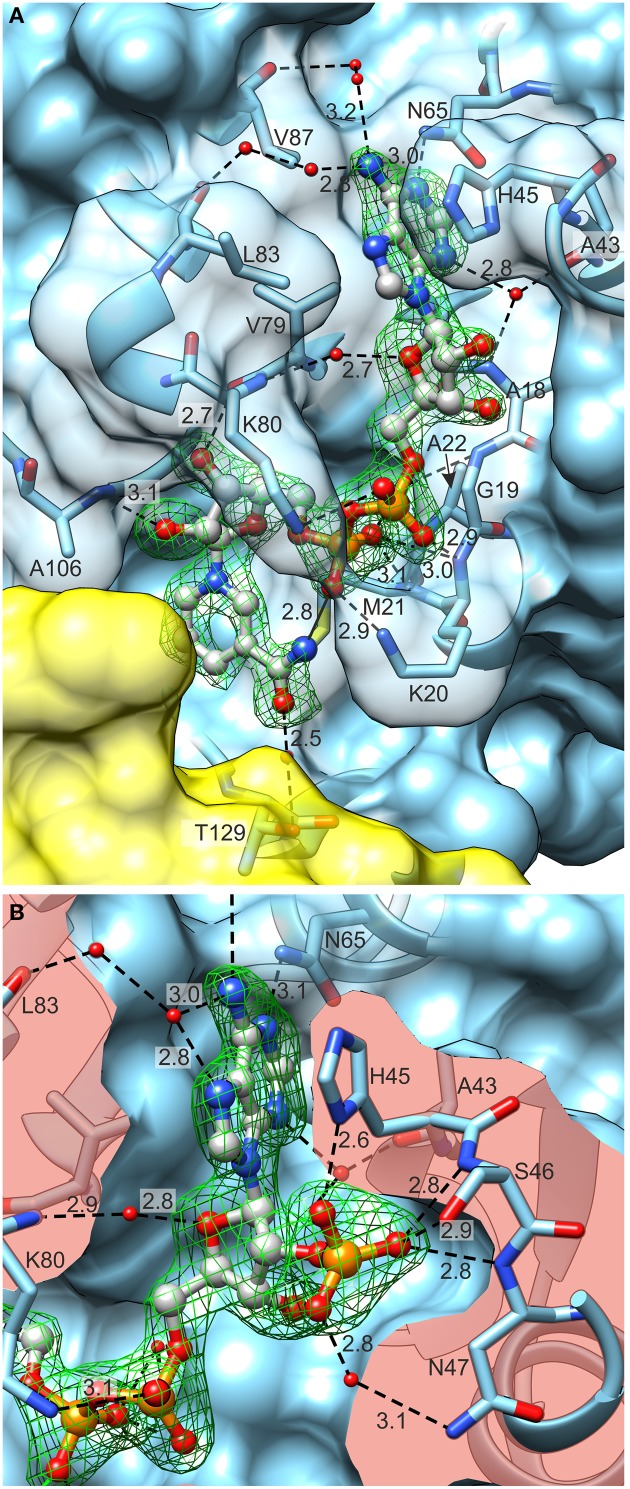
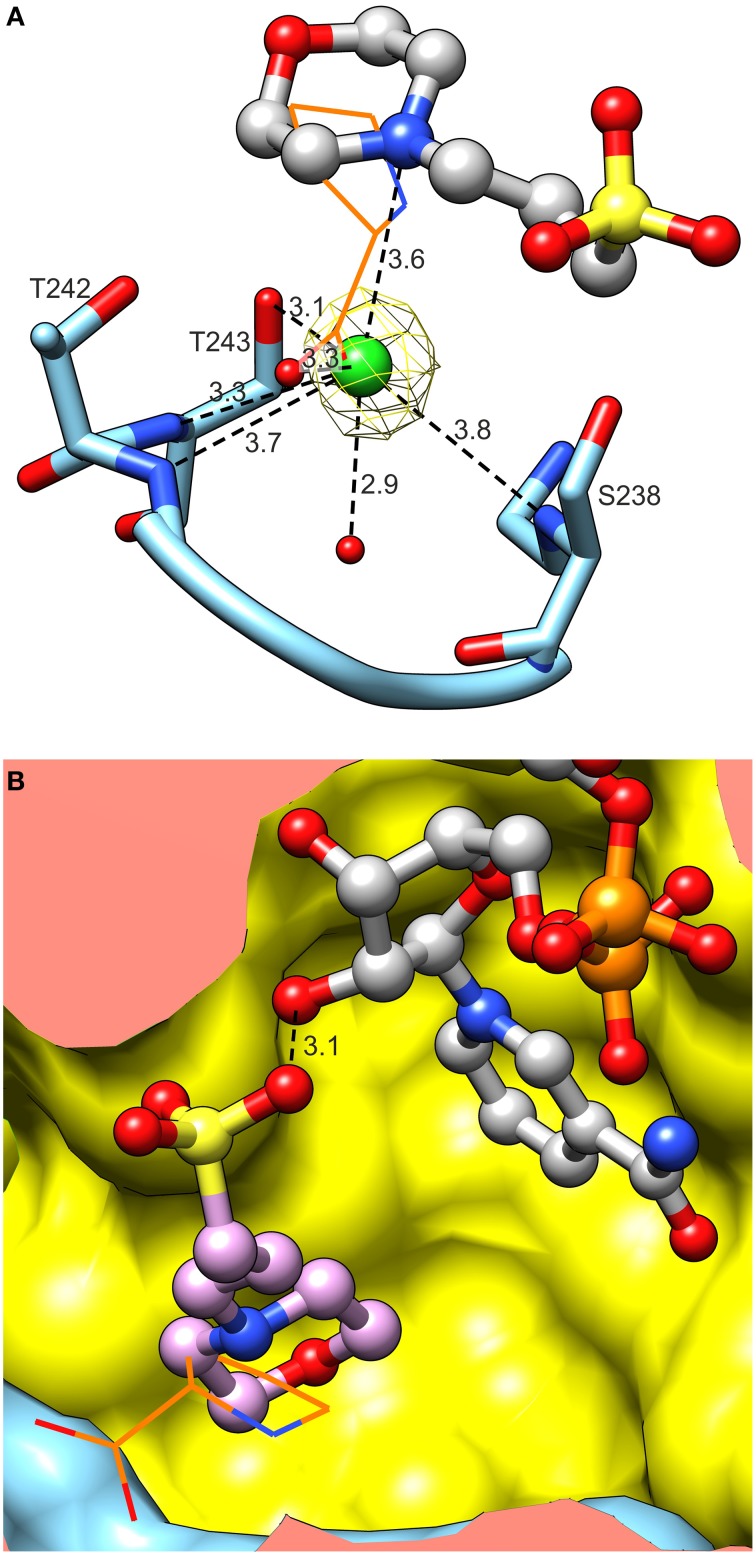
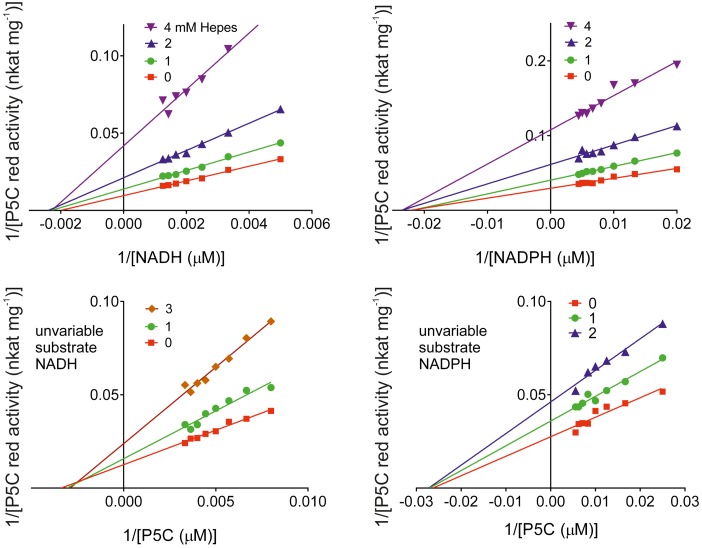
The two pathways for proline biosynthesis in higher plants share the last step, the conversion of δ(1)-pyrroline-5-carboxylate (P5C) to L-proline, which is catalyzed by P5C reductase (P5CR, EC 1.5.1.2) with the use of NAD(P)H as a coenzyme. There is increasing amount of evidence to suggest a complex regulation of P5CR activity at the post-translational level, yet the molecular basis of these mechanisms is unknown. Here we report the three-dimensional structure of the P5CR enzyme from the model legume Medicago truncatula (Mt). The crystal structures of unliganded MtP5CR decamer, and its complexes with the products NAD(+), NADP(+), and L-proline were refined using x-ray diffraction data (at 1.7, 1.85, 1.95, and 2.1 Å resolution, respectively). Based on the presented structural data, the coenzyme preference for NADPH over NADH was explained, and NADPH is suggested to be the only coenzyme used by MtP5CR in vivo. Furthermore, the insensitivity of MtP5CR to feed-back inhibition by proline, revealed by enzymatic analysis, was correlated with structural features. Additionally, a mechanism for the modulation of enzyme activity by chloride anions is discussed, as well as the rationale for the possible development of effective enzyme inhibitors.
Keywords: P5C reductase; P5CR; abiotic stress; coenzyme preference; decamer; protein structure; salt stress.
Figures

Similar articles
-
Δ1-Pyrroline-5-carboxylate reductase from Arabidopsis thaliana: stimulation or inhibition by chloride ions and feedback regulation by proline depend on whether NADPH or NADH acts as co-substrate.New Phytol. 2014 May;202(3):911-919. doi: 10.1111/nph.12701. Epub 2014 Jan 28. New Phytol. 2014. PMID: 24467670
-
Crystal structures of delta1-pyrroline-5-carboxylate reductase from human pathogens Neisseria meningitides and Streptococcus pyogenes.J Mol Biol. 2005 Nov 18;354(1):91-106. doi: 10.1016/j.jmb.2005.08.036. Epub 2005 Sep 2. J Mol Biol. 2005. PMID: 16233902 Free PMC article.
-
Functional Characterization of Saccharomyces cerevisiae P5C Reductase, the Enzyme at the Converging Point of Proline and Arginine Metabolism.Microorganisms. 2022 Oct 20;10(10):2077. doi: 10.3390/microorganisms10102077. Microorganisms. 2022. PMID: 36296354 Free PMC article.
-
Regulation of levels of proline as an osmolyte in plants under water stress.Plant Cell Physiol. 1997 Oct;38(10):1095-102. doi: 10.1093/oxfordjournals.pcp.a029093. Plant Cell Physiol. 1997. PMID: 9399433 Review.
-
Isozymes of P5C reductase (PYCR) in human diseases: focus on cancer.Amino Acids. 2021 Dec;53(12):1835-1840. doi: 10.1007/s00726-021-03048-x. Epub 2021 Jul 22. Amino Acids. 2021. PMID: 34291342 Review.
Cited by
-
Proline Tagging for Stress Tolerance in Plants.Int J Genomics. 2025 Apr 2;2025:9348557. doi: 10.1155/ijog/9348557. eCollection 2025. Int J Genomics. 2025. PMID: 40207093 Free PMC article. Review.
-
Phenyl-substituted aminomethylene-bisphosphonates inhibit human P5C reductase and show antiproliferative activity against proline-hyperproducing tumour cells.J Enzyme Inhib Med Chem. 2021 Dec;36(1):1248-1257. doi: 10.1080/14756366.2021.1919890. J Enzyme Inhib Med Chem. 2021. PMID: 34107832 Free PMC article.
-
Structure, biochemistry, and gene expression patterns of the proline biosynthetic enzyme pyrroline-5-carboxylate reductase (PYCR), an emerging cancer therapy target.Amino Acids. 2021 Dec;53(12):1817-1834. doi: 10.1007/s00726-021-02999-5. Epub 2021 May 18. Amino Acids. 2021. PMID: 34003320 Free PMC article. Review.
-
Enzymology and Regulation of δ1-Pyrroline-5-Carboxylate Synthetase 2 From Rice.Front Plant Sci. 2021 Sep 14;12:672702. doi: 10.3389/fpls.2021.672702. eCollection 2021. Front Plant Sci. 2021. PMID: 34603346 Free PMC article.
-
Exogenous acetone O-(4-chlorophenylsulfonyl) oxime alleviates Cd stress-induced photosynthetic damage and oxidative stress by regulating the antioxidant defense mechanism in Zea mays.Physiol Mol Biol Plants. 2022 Dec;28(11-12):2069-2083. doi: 10.1007/s12298-022-01258-5. Epub 2022 Dec 3. Physiol Mol Biol Plants. 2022. PMID: 36573151 Free PMC article.
References
-
- Choudhary N. L., Sairam R. K., Tyagi A. (2005). Expression of δ1-pyrroline-5-carboxylate synthetase gene during drought in rice (Oryza sativa L.). Indian J. Biochem. Biophys. 42, 366–370. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources