

Review
doi: 10.1038/nsmb.3092.
Complex interactions between the DNA-damage response and mammalian telomeres
Affiliations
- PMID: 26581520
- PMCID: PMC4739752
- DOI: 10.1038/nsmb.3092
Item in Clipboard
Review
Complex interactions between the DNA-damage response and mammalian telomeres
Nat Struct Mol Biol.
2015 Nov.
Abstract
Natural chromosome ends resemble double-stranded DNA breaks, but they do not activate a damage response in healthy cells. Telomeres therefore have evolved to solve the 'end-protection problem' by inhibiting multiple DNA damage-response pathways. During the past decade, the view of telomeres has progressed from simple caps that hide chromosome ends to complex machineries that have an active role in organizing the genome. Here we focus on mammalian telomeres and summarize and interpret recent discoveries in detail, focusing on how repair pathways are inhibited, how resection and replication are controlled and how these mechanisms govern cell fate during senescence, crisis and transformation.
Figures

Repression of DNA-damage signaling pathways at telomeres. (a) ATM kinase pathway. (b) ATR kinase pathway. (c) Alternative NHEJ repair pathway. (d) Classical NHEJ repair pathway. (e) Homologous-recombination pathway. Steps of DNA damage–repair inhibition by shelterin or accessory factors are highlighted by red circles. (1) TRF2, potentially through its TRFH domain, and supported by Rap1, forms the t loop, a DNA structure that hides chromosome ends from the MRN–ATM factors–,. (2) The t loop also prevents loading of the Ku heterodimer on chromosome ends. (3) TRF2iDDR inhibits the recruitment of RNF168 at telomeres. (4) TRF2 interacts with the helix α5 domain of Ku70, thereby blocking its heterotetramerization. (5) Rap1 has been proposed to directly block c-NHEJ. (6) TPP1–POT1 prevents ATR activation, most probably by preventing binding of RPA on the overhang and on the ssTTAGGG at stalled replication forks–,. (7) The Ku heterodimer, TRF2 and, more importantly, TPP1–POT1 redundantly repress alt-NHEJ at telomeres,,. (8) The shelterin factors Rap1 and POT1 as well as Ku70–Ku80 are necessary to block HR between sister telomeres,,,. (9) The N-terminal basic domain of TRF2 blocks NBS1–XRCC3–mediated recombination events at the D loop that would lead to t-loop excision. Ku depletion induces t-loop excision as well–. (10) At deprotected telomeres, 53BP1, RIF1 and MAD2L2 protect chromosome ends from BRCA1–CtIP–EXO1–mediated 5′ resection–. LIG, ligase; Pol, polymerase; p, phospho-.

Functions and interactions of TRF2 domains. TRF2 possesses four distinct domains, the N-terminal basic domain,,, the TRFH domain10,44,46,96–100, the hinge domain10,14,93 and the MYB DNA-binding domain,. Human (H) or mouse (M) amino acid residues that are involved in protein-protein interactions or that disrupt interactions when mutated are specified in the third column.
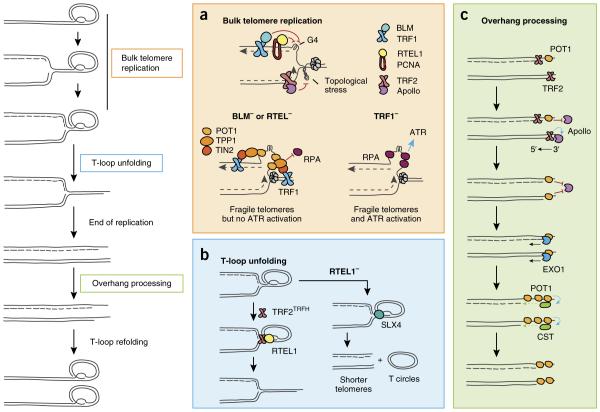
Major DNA-repair factors involved in telomere maintenance during S phase. Replication initiates within subtelomeres and progresses until it encounters the D loop, which must be dismantled to allow replication to be completed. The overhangs are processed on both sister chromatids, and the t loop is refolded. (a) TRF1 and proliferating cell nuclear antigen (PCNA) recruit BLM and RTEL1, both of which unfold G-quadruplex structures on the lagging strand,, while Apollo recruitment by TRF2 prevents accumulation of topological stress ahead of the fork. Without BLM or RTEL1, the replication fork is blocked, thus leading to G-rich ssDNA accumulation. A bridge consisting of TRF1, TIN2 and TPP1 anchors POT1 on this ssDNA, preventing RPA loading and ATR activation,. (b) Once the bulk telomere is replicated, TRF2TRFH recruits RTEL1, which unfolds the t loop. In the absence of RTEL1, t loops are excised by SLX4, thus leading to telomere shortening and t-circle accumulation,. (c) Unidirectional replication results in blunt ends at the daughter chromatid formed by leading-strand replication, whereas lagging-strand replication leaves a short overhang that is bound by POT1. After termination of replication, TRF2 recruits Apollo to the leading-strand chromatid, and Apollo resects the 5′ end and generates a short overhang. Binding of POT1 blocks further resection by Apollo. A second round of overhang processing occurs during late S-G2, when EXO1 further resects the 5′ ends of both sister chromatids. Finally, the CST complex is recruited to the extended overhangs to fill in the lagging strand and reduce overhang length–.

Replicative senescence and crisis are two proliferative barriers controlled by telomere deprotection. Young fibroblasts contain long, fully protected telomeres (blue), which erode over the course of cellular division. Some telomeres become dysfunctional and enter the intermediate state of deprotection (orange), which is characterized by activation of the DNA-damage response, yet retain enough protective shelterin to inhibit fusions,. Telomere shortening and deprotection increase with population doublings until five or more dysfunctional telomeres are encountered, a damage threshold that activates p53 and leads to entry into replicative senescence. Upon loss of p53 and retinoblastoma protein (Rb), cells bypass senescence and continue to grow. Telomeres continue to shorten until they become too short to retain any protective properties. Uncapped telomeres (red) fuse, thus leading to prolonged mitotic arrest that amplifies telomere deprotection and causes cell death in crisis,.
References
-
- de Lange T. Shelterin: the protein complex that shapes and safeguards human telomeres. Genes Dev. 2005;19:2100–2110. - PubMed
-
- Karlseder J. p53- and ATM-dependent apoptosis induced by telomeres lacking TRF2. Science. 1999;283:1321–1325. - PubMed
-
- Smogorzewska A, Karlseder J, Holtgreve-Grez H, Jauch A, de Lange T. DNA ligase IV-dependent NHEJ of deprotected mammalian telomeres in G1 and G2. Curr. Biol. 2002;12:1635–1644. - PubMed
-
- van Steensel B, Smogorzewska A, de Lange T. TRF2 protects human telomeres from end-to-end fusions. Cell. 1998;92:401–413. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
