

Macroevolutionary assembly of ant/plant symbioses: Pseudomyrmex ants and their ant-housing plants in the Neotropics
- PMID: 26582029
- PMCID: PMC4685824
- DOI: 10.1098/rspb.2015.2200
Macroevolutionary assembly of ant/plant symbioses: Pseudomyrmex ants and their ant-housing plants in the Neotropics
Abstract
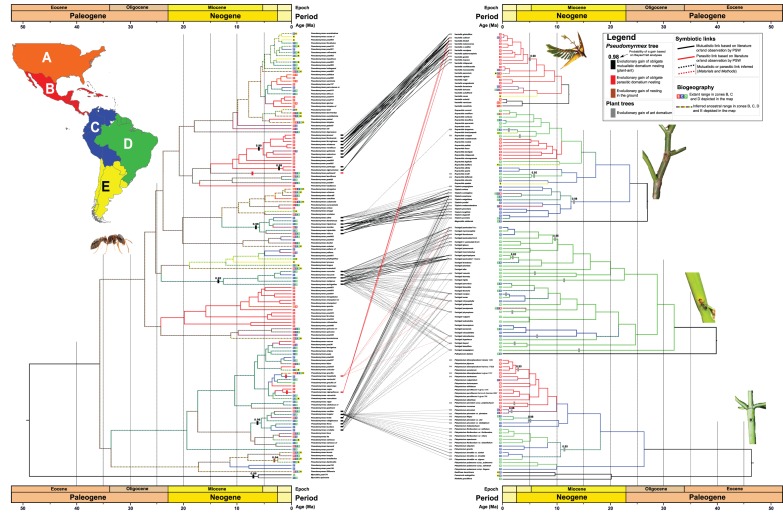
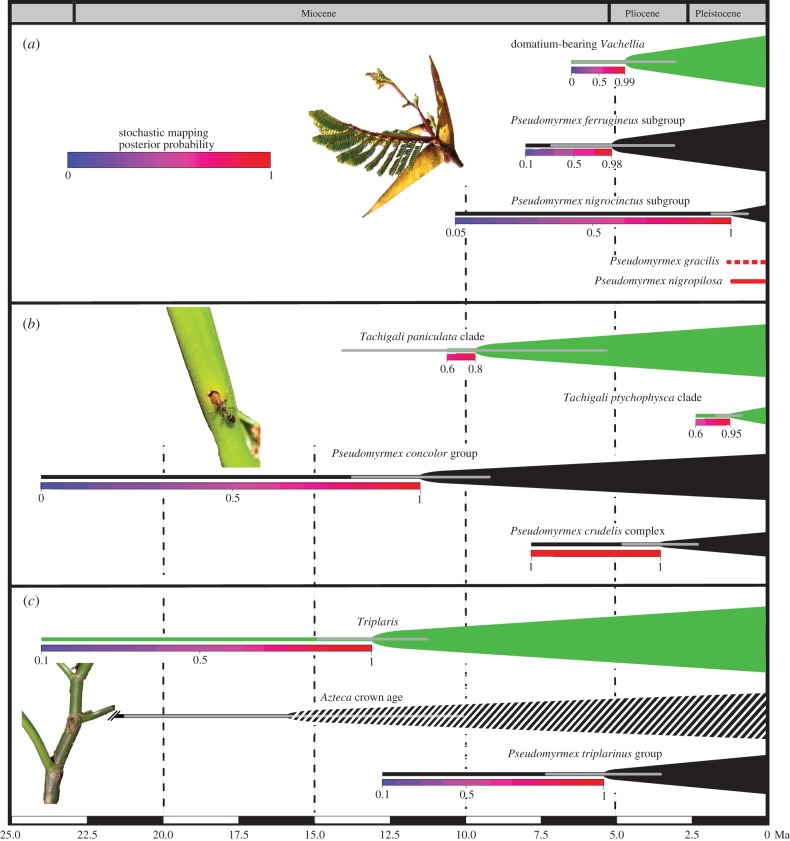
Symbioses include some of the clearest cases of coevolution, but their origin, loss or reassembly with different partners can rarely be inferred. Here we use ant/plant symbioses involving three plant clades to investigate the evolution of symbioses. We generated phylogenies for the big-eyed arboreal ants (Pseudomyrmecinae), including 72% of their 286 species, as well as for five of their plant host groups, in each case sampling more than 61% of the species. We show that the ant-housing Vachellia (Mimosoideae) clade and its ants co-diversified for the past 5 Ma, with some species additionally colonized by younger plant-nesting ant species, some parasitic. An apparent co-radiation of ants and Tachigali (Caesalpinioideae) was followed by waves of colonization by the same ant clade, and subsequent occupation by a younger ant group. Wide crown and stem age differences between the ant-housing genus Triplaris (Polygonaceae) and its obligate ant inhabitants, and stochastic trait mapping, indicate that its domatium evolved earlier than the ants now occupying it, suggesting previous symbioses that dissolved. Parasitic ant species evolved from generalists, not from mutualists, and are younger than the mutualistic systems they parasitize. Our study illuminates the macroevolutionary assembly of ant/plant symbioses, which has been highly dynamic, even in very specialized systems.
Keywords: Pseudomyrmex; ant/plant coevolution; co-radiation; molecular clocks; secondary colonization; symbiosis.
© 2015 The Author(s).
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous