

Two Na+ Sites Control Conformational Change in a Neurotransmitter Transporter Homolog
- PMID: 26582198
- PMCID: PMC4714228
- DOI: 10.1074/jbc.M115.692012
Two Na+ Sites Control Conformational Change in a Neurotransmitter Transporter Homolog
Abstract
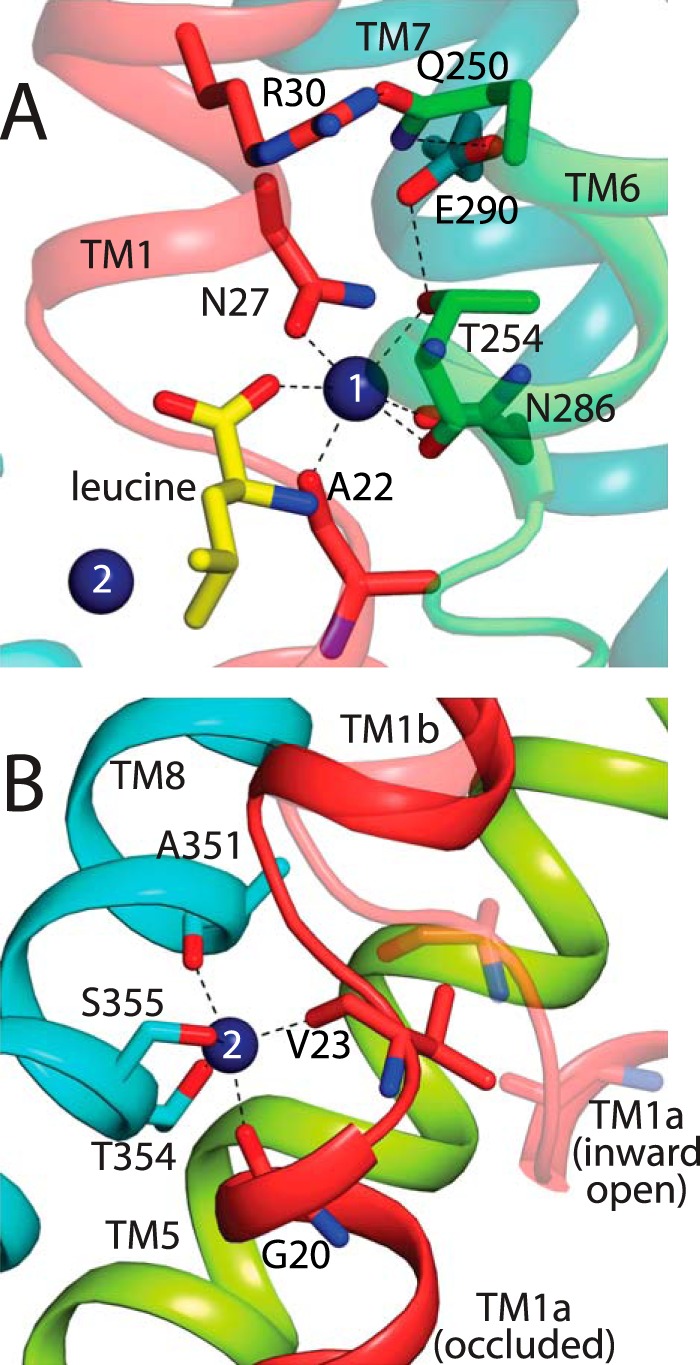
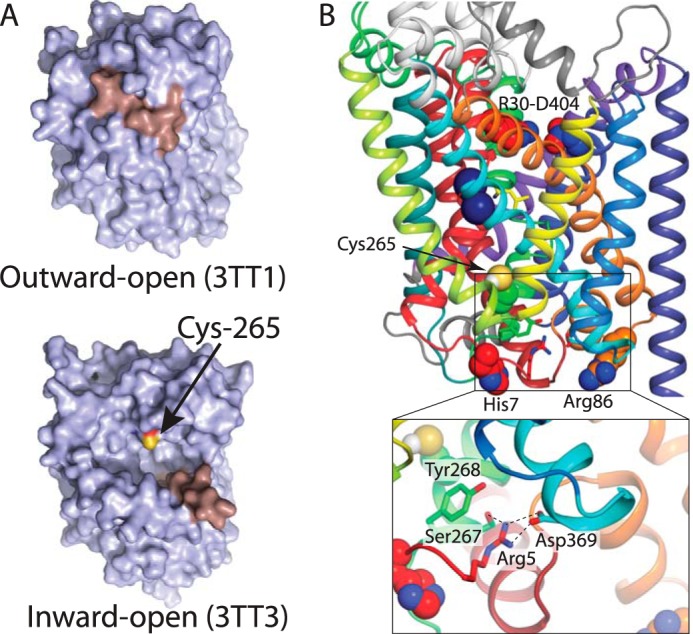
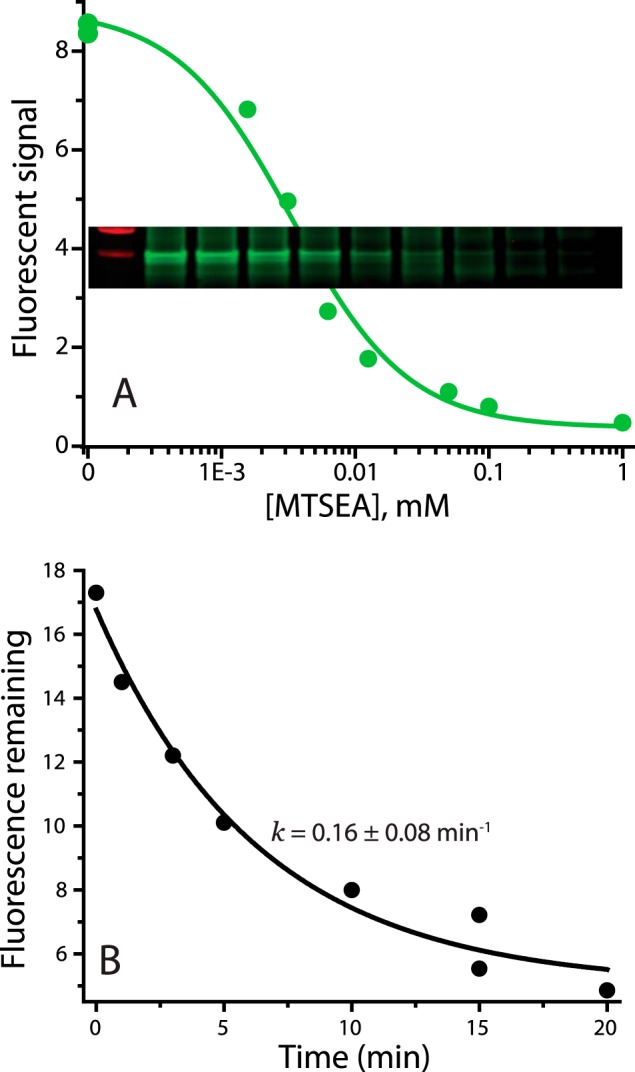
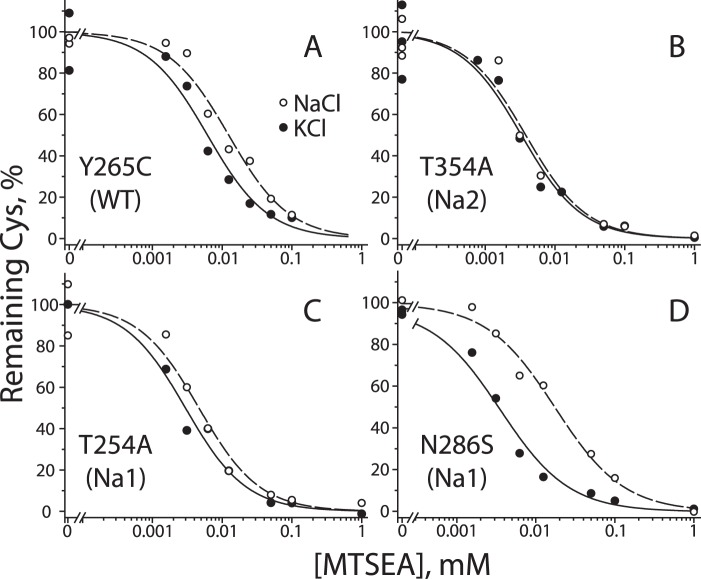
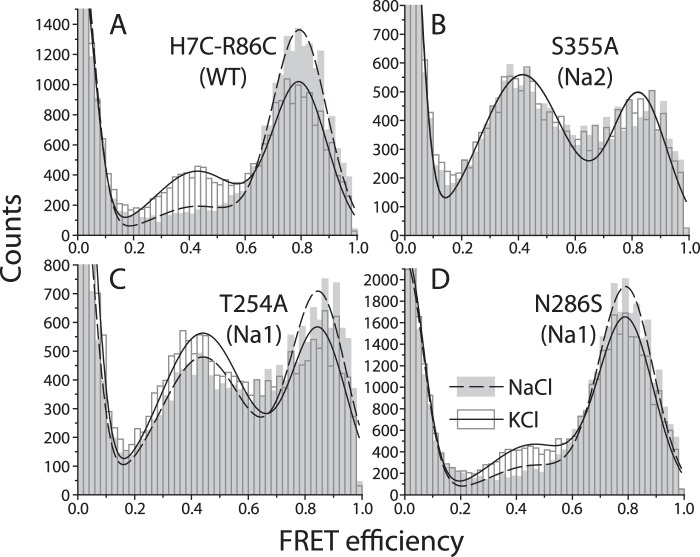
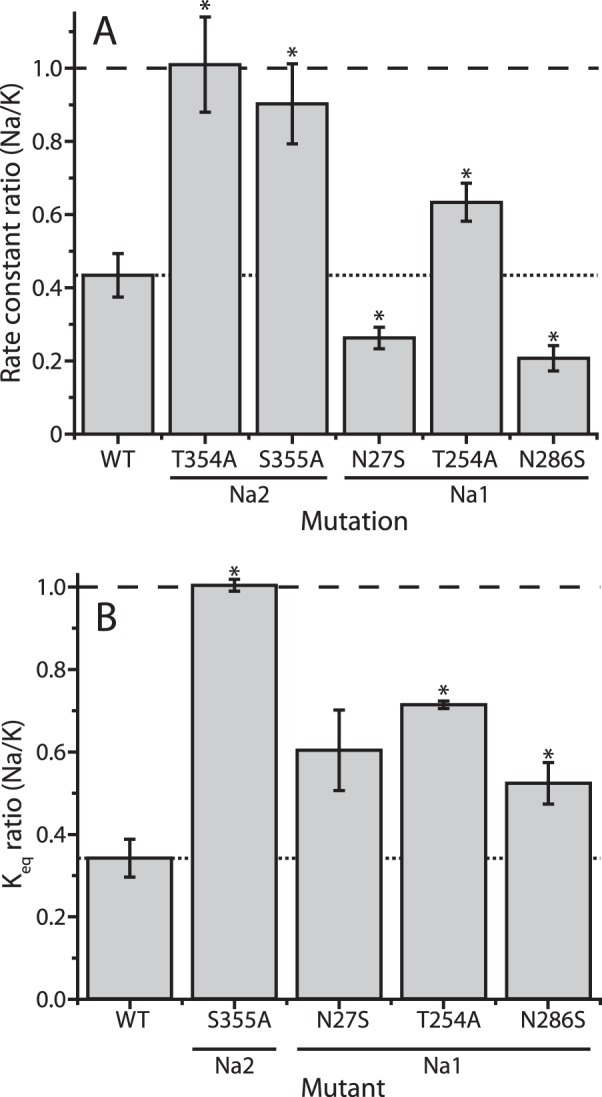
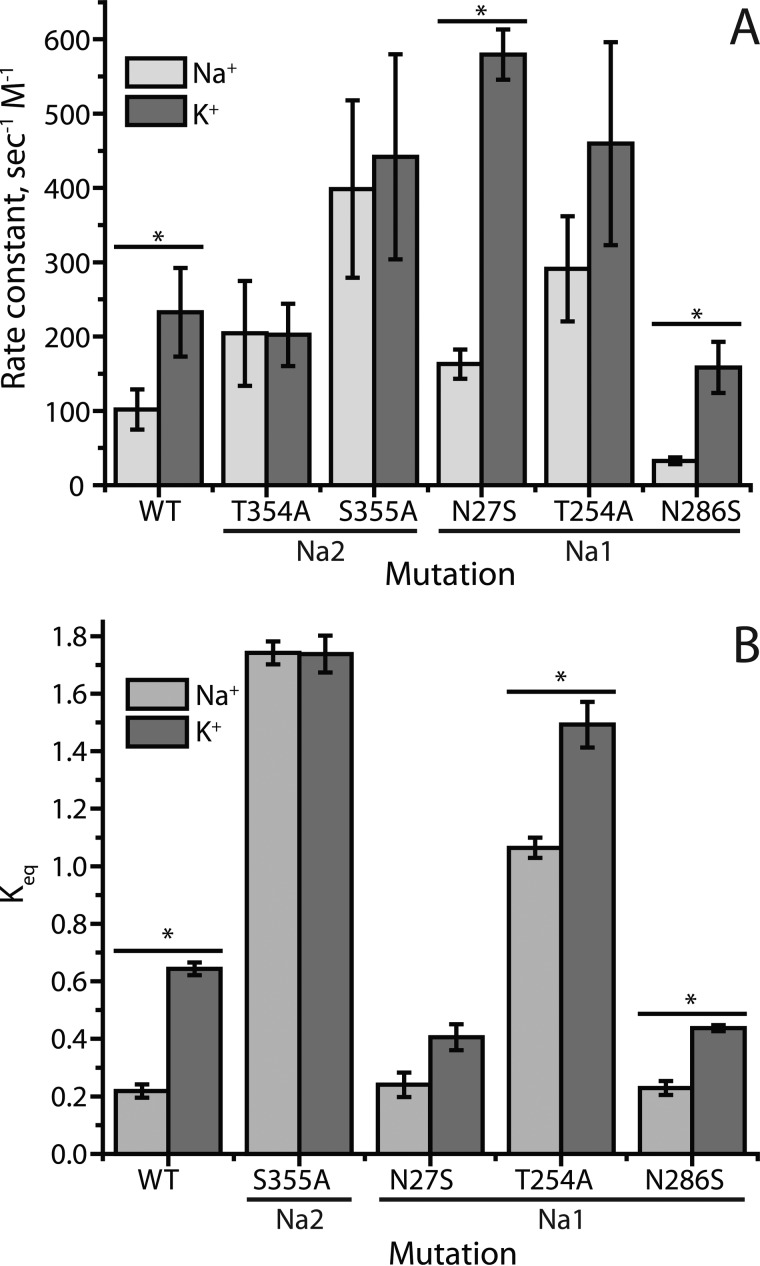
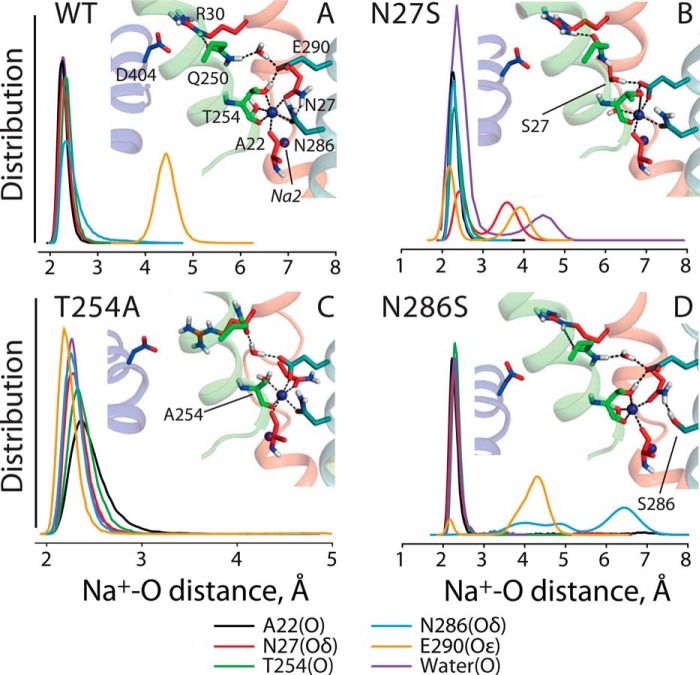
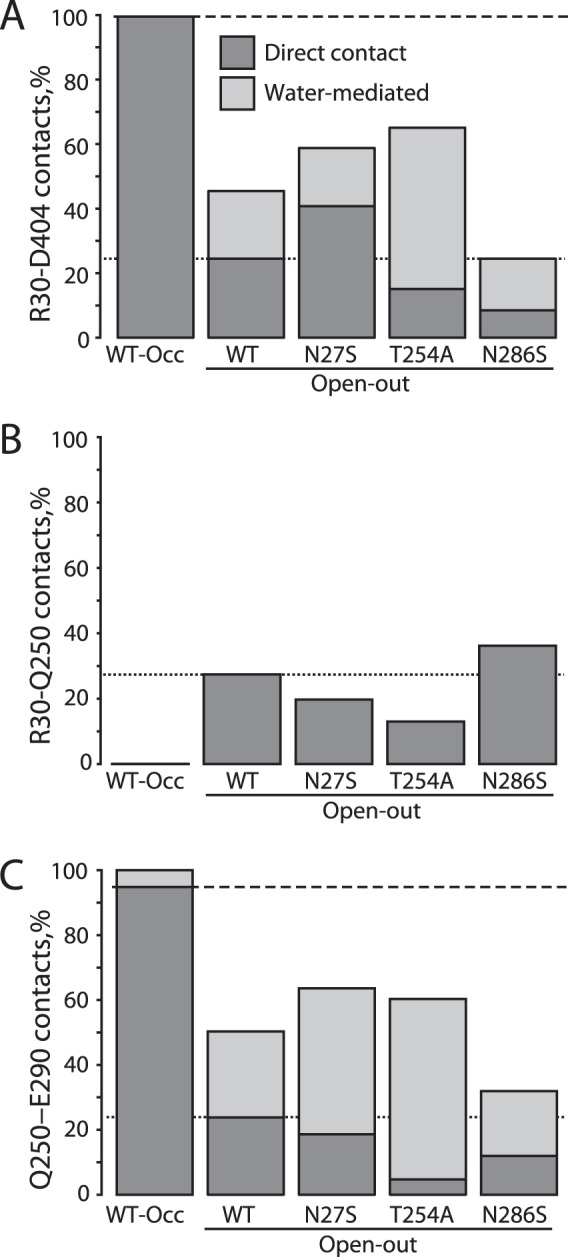
In LeuT, a prokaryotic homolog of neurotransmitter transporters, Na(+) stabilizes outward-open conformational states. We examined how each of the two LeuT Na(+) binding sites contributes to Na(+)-dependent closure of the cytoplasmic pathway using biochemical and biophysical assays of conformation. Mutating either of two residues that contribute to the Na2 site completely prevented cytoplasmic closure in response to Na(+), suggesting that Na2 is essential for this conformational change, whereas Na1 mutants retained Na(+) responsiveness. However, mutation of Na1 residues also influenced the Na(+)-dependent conformational change in ways that varied depending on the position mutated. Computational analyses suggest those mutants influence the ability of Na1 binding to hydrate the substrate pathway and perturb an interaction network leading to the extracellular gate. Overall, the results demonstrate that occupation of Na2 stabilizes outward-facing conformations presumably through a direct interaction between Na(+) and transmembrane helices 1 and 8, whereas Na(+) binding at Na1 influences conformational change through a network of intermediary interactions. The results also provide evidence that N-terminal release and helix motions represent distinct steps in cytoplasmic pathway opening.
Keywords: alternating access; conformational change; coupling; membrane transport; molecular dynamics; neurotransmitter; single-molecule biophysics; sodium; transport.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Quick M., Yano H., Goldberg N. R., Duan L., Beuming T., Shi L., Weinstein H., and Javitch J. A. (2006) State-dependent conformations of the translocation pathway in the tyrosine transporter Tyt1, a novel neurotransmitter:sodium symporter from Fusobacterium nucleatum. J. Biol. Chem. 281, 26444–26454 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
