

A Single Glycan at the 99-Loop of Human Kallikrein-related Peptidase 2 Regulates Activation and Enzymatic Activity
- PMID: 26582203
- PMCID: PMC4705380
- DOI: 10.1074/jbc.M115.691097
A Single Glycan at the 99-Loop of Human Kallikrein-related Peptidase 2 Regulates Activation and Enzymatic Activity
Abstract
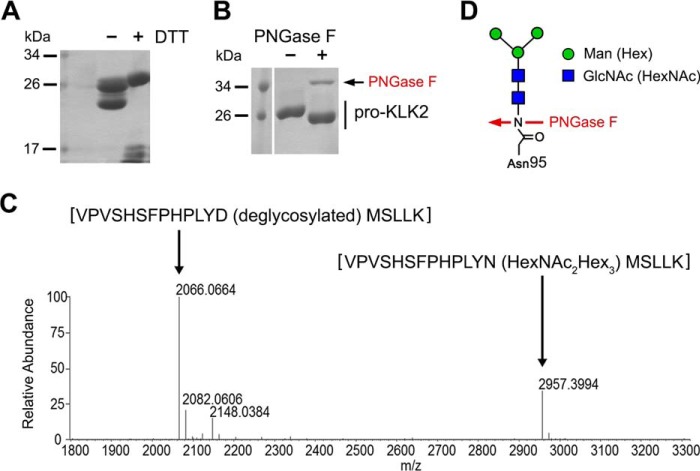
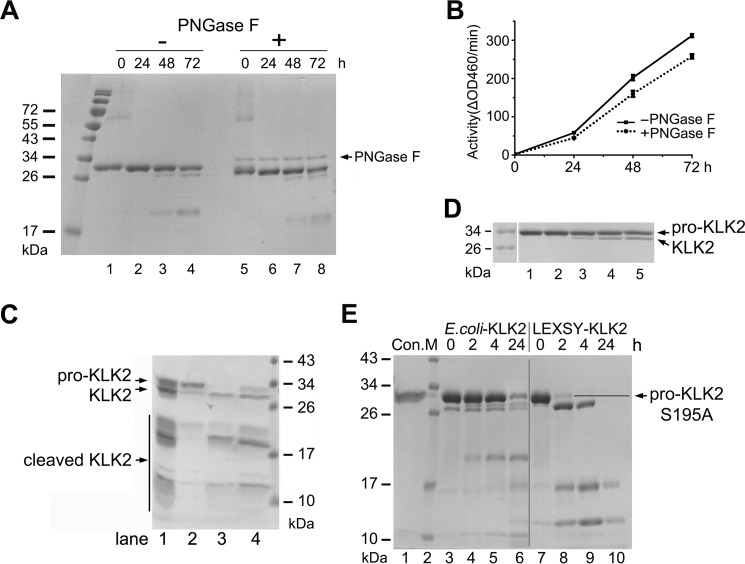
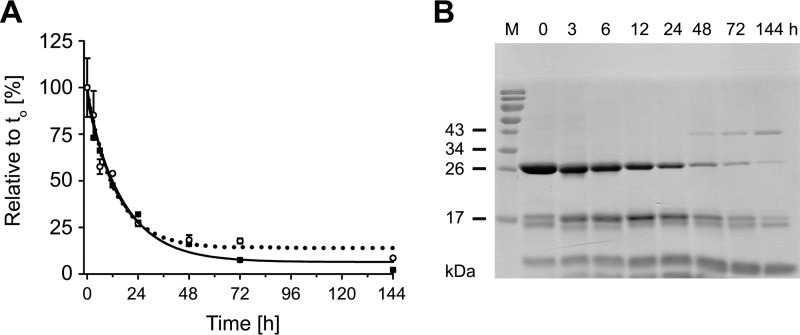
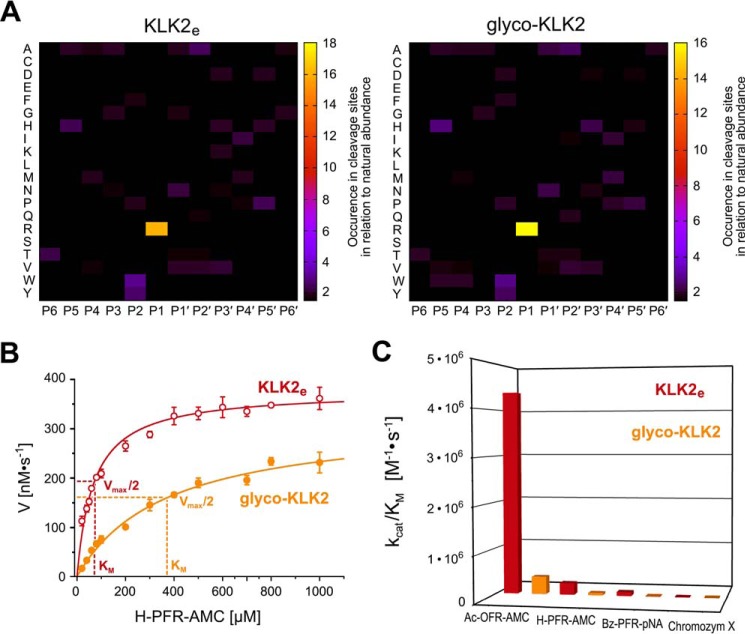
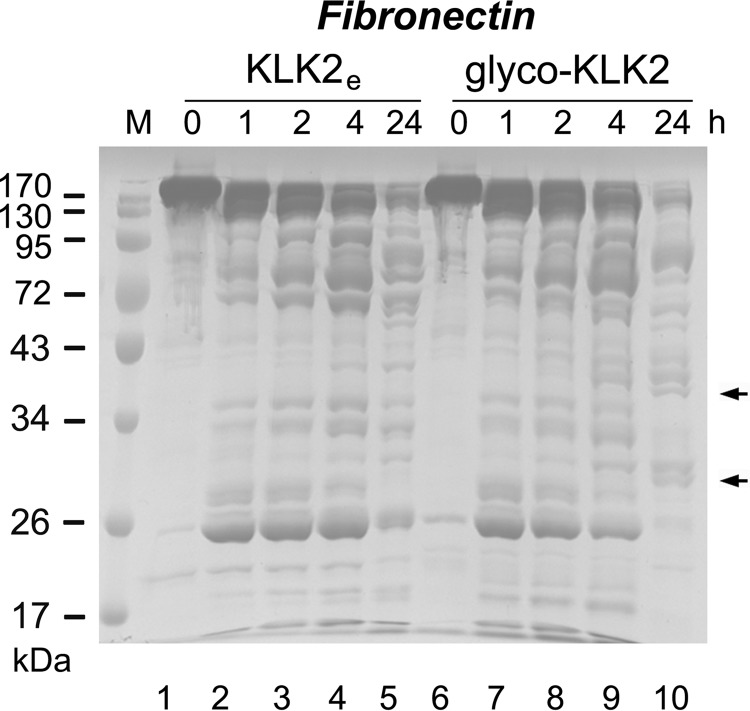
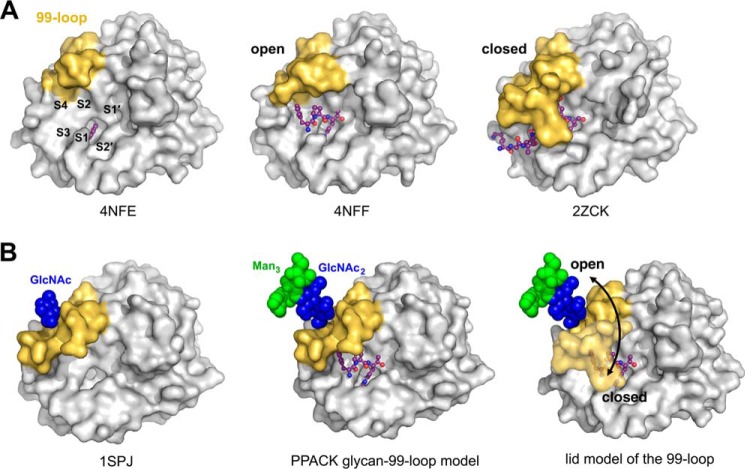
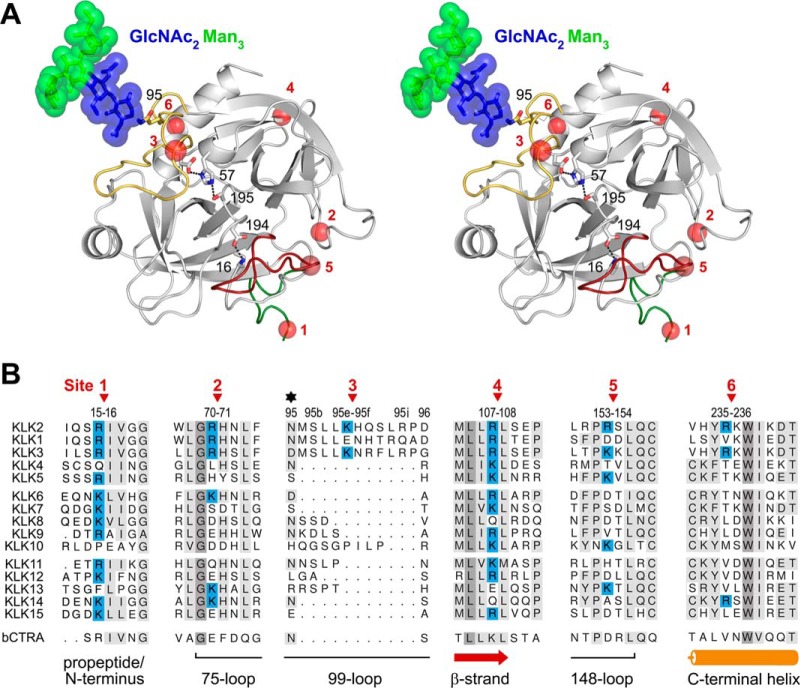
Human kallikrein-related peptidase 2 (KLK2) is a key serine protease in semen liquefaction and prostate cancer together with KLK3/prostate-specific antigen. In order to decipher the function of its potential N-glycosylation site, we produced pro-KLK2 in Leishmania tarentolae cells and compared it with its non-glycosylated counterpart from Escherichia coli expression. Mass spectrometry revealed that Asn-95 carries a core glycan, consisting of two GlcNAc and three hexoses. Autocatalytic activation was retarded in glyco-pro-KLK2, whereas the activated glyco-form exhibited an increased proteolytic resistance. The specificity patterns obtained by the PICS (proteomic identification of protease cleavage sites) method are similar for both KLK2 variants, with a major preference for P1-Arg. However, glycosylation changes the enzymatic activity of KLK2 in a drastically substrate-dependent manner. Although glyco-KLK2 has a considerably lower catalytic efficiency than glycan-free KLK2 toward peptidic substrates with P2-Phe, the situation was reverted toward protein substrates, such as glyco-pro-KLK2 itself. These findings can be rationalized by the glycan-carrying 99-loop that prefers to cover the active site like a lid. By contrast, the non-glycosylated 99-loop seems to favor a wide open conformation, which mostly increases the apparent affinity for the substrates (i.e. by a reduction of Km). Also, the cleavage pattern and kinetics in autolytic inactivation of both KLK2 variants can be explained by a shift of the target sites due to the presence of the glycan. These striking effects of glycosylation pave the way to a deeper understanding of kallikrein-related peptidase biology and pathology.
Keywords: N-linked glycosylation; autolytic inactivation; enzyme kinetics; kallikrein; prostate cancer; serine protease; substrate specificity; zymogen activation.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Raspi G. (1996) Kallikrein and kallikrein-like proteinases: purification and determination by chromatographic and electrophoretic methods. J. Chromatogr. B Biomed. Appl. 684, 265–287 - PubMed
-
- Avgeris M., Mavridis K., and Scorilas A. (2012) Kallikrein-related peptidases in prostate, breast, and ovarian cancers: from pathobiology to clinical relevance. Biol. Chem. 393, 301–317 - PubMed
-
- Fukushima D., Kitamura N., and Nakanishi S. (1985) Nucleotide sequence of cloned cDNA for human pancreatic kallikrein. Biochemistry 24, 8037–8043 - PubMed
-
- Baker A. R., and Shine J. (1985) Human-kidney kallikrein: cDNA cloning and sequence-analysis. DNA 4, 445–450 - PubMed
-
- Schedlich L. J., Bennetts B. H., and Morris B. J. (1987) Primary structure of a human glandular kallikrein gene. DNA 6, 429–437 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Miscellaneous
