

Smad4 represses the generation of memory-precursor effector T cells but is required for the differentiation of central memory T cells
- PMID: 26583325
- PMCID: PMC4670941
- DOI: 10.1038/cddis.2015.337
Smad4 represses the generation of memory-precursor effector T cells but is required for the differentiation of central memory T cells
Abstract
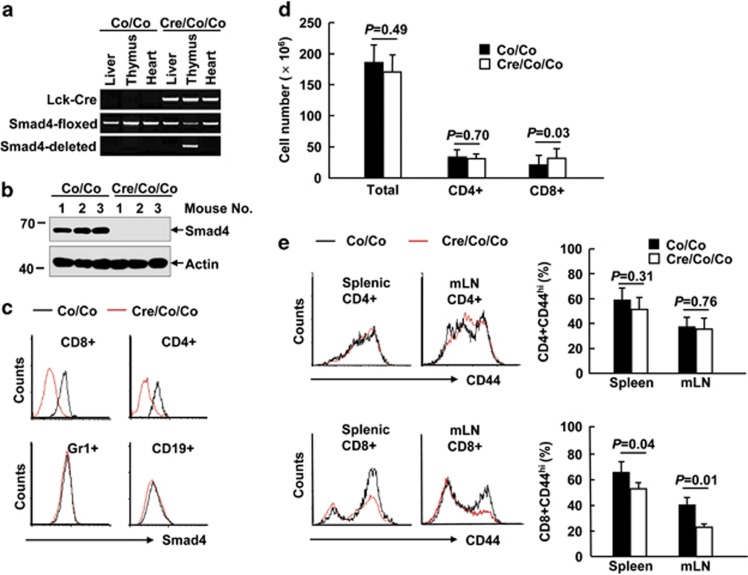
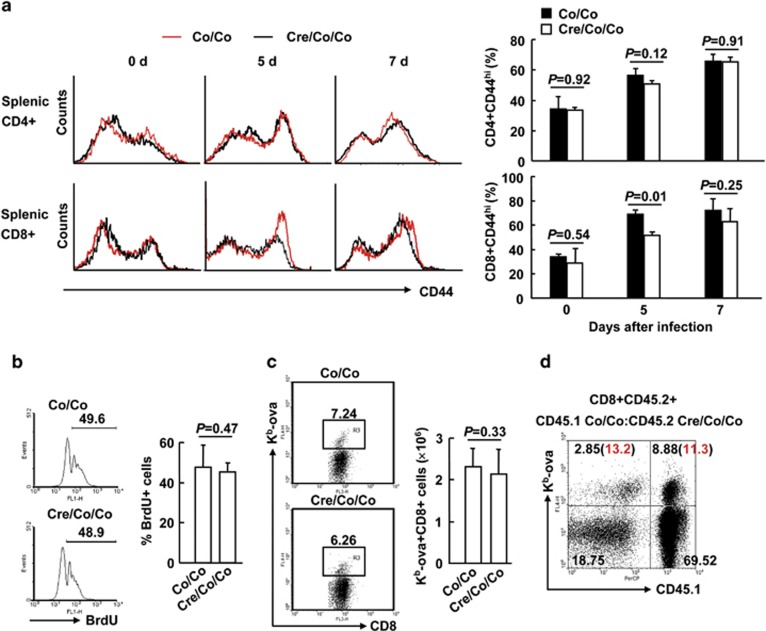
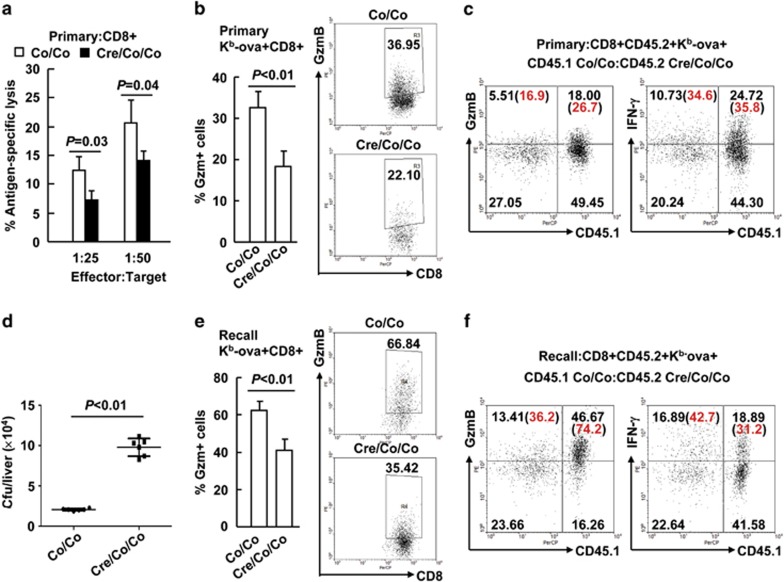
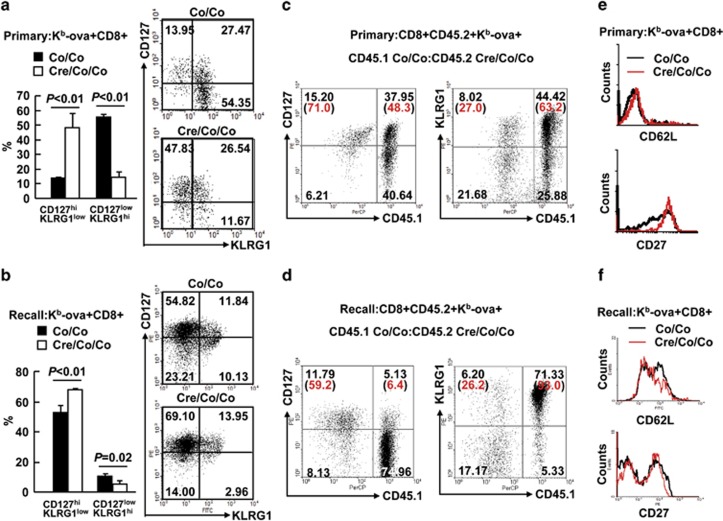
The transcriptional regulation underlying the differentiation of CD8(+) effector and memory T cells remains elusive. Here, we show that 18-month-old mice lacking the transcription factor Smad4 (homolog 4 of mothers against decapentaplegic, Drosophila), a key intracellular signaling effector for the TGF-β superfamily, in T cells exhibited lower percentages of CD44(hi)CD8(+) T cells. To explore the role of Smad4 in the activation/memory of CD8(+) T cells, 6- to 8-week-old mice with or without Smad4 in T cells were challenged with Listeria monocytogenes. Smad4 deficiency did not affect antigen-specific CD8(+) T-cell expansion but led to partially impaired cytotoxic function. Less short-lived effector T cells but more memory-precursor effector T cells were generated in the absence of Smad4. Despite that, Smad4 deficiency led to reduced memory CD8(+) T-cell responses. Further exploration revealed that the generation of central memory T cells was impaired in the absence of Smad4 and the cells showed survival issue. In mechanism, Smad4 deficiency led to aberrant transcriptional programs in antigen-specific CD8(+) T cells. These findings demonstrated an essential role of Smad4 in the control of effector and memory CD8(+) T-cell responses to infection.
Figures
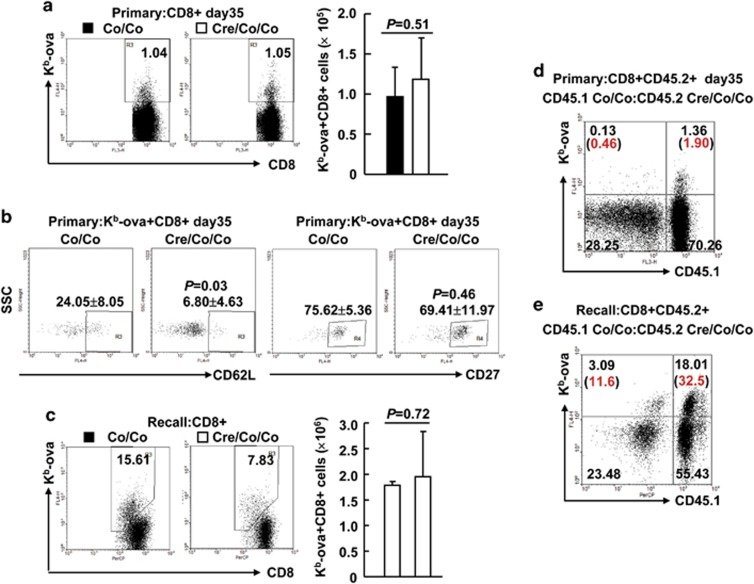
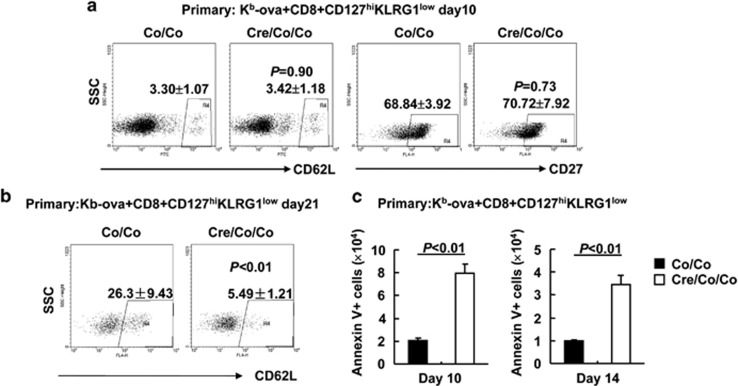
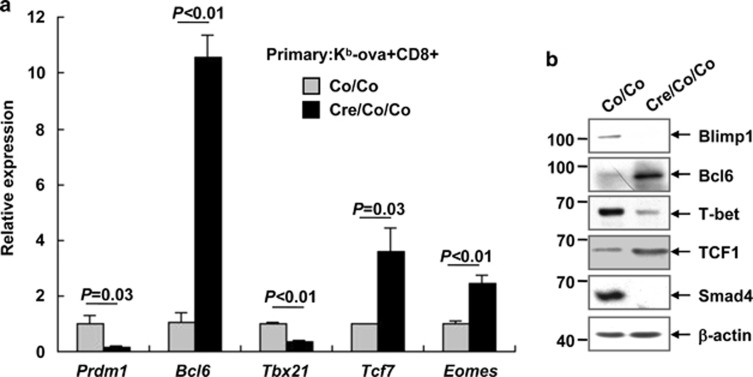
References
-
- 1Klenerman P, Hill A. T cells and viral persistence: lessons from diverse infections. Nat Immunol 2005; 6: 873–879. - PubMed
-
- 3Kaech SM, Tan JT, Wherry EJ, Konieczny BT, Surh CD, Ahmed R. Selective expression of the interleukin 7 receptor identifies effector CD8 T cells that give rise to long-lived memory cells. Nat Immunol 2003; 4: 1191–1198. - PubMed
-
- 5Intlekofer AM, Takemoto N, Wherry EJ, Longworth SA, Northrup JT, Palanivel VR et al. Effector and memory CD8+ T cell fate coupled by T-bet and eomesodermin. Nat Immunol 2005; 6: 1236–1244. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
