

High telomerase is a hallmark of undifferentiated spermatogonia and is required for maintenance of male germline stem cells
- PMID: 26584619
- PMCID: PMC4691947
- DOI: 10.1101/gad.271783.115
High telomerase is a hallmark of undifferentiated spermatogonia and is required for maintenance of male germline stem cells
Abstract
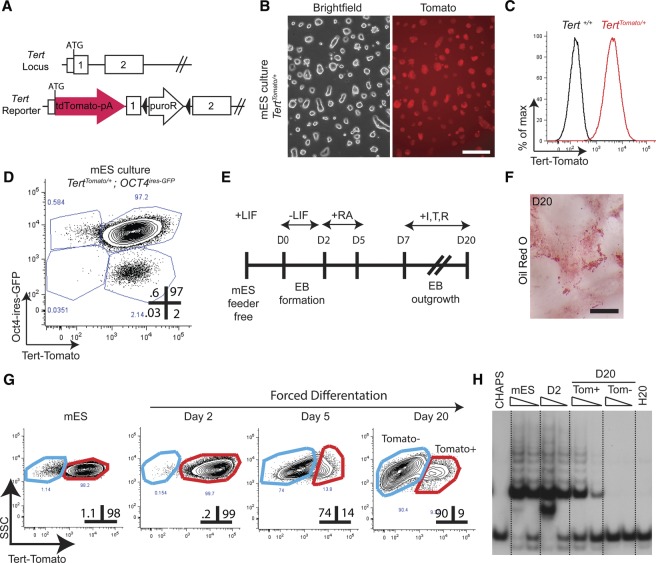
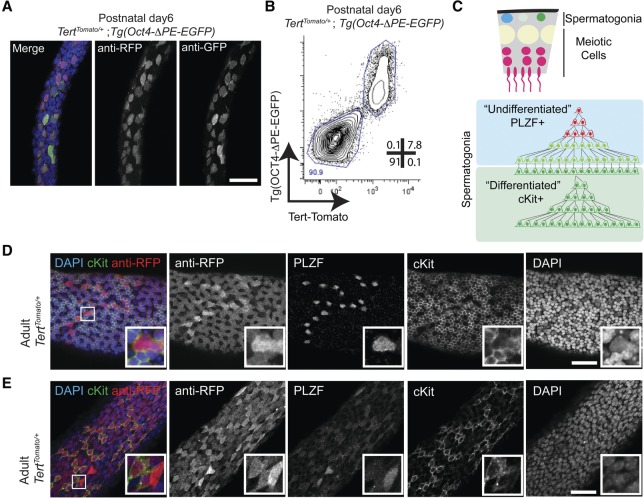
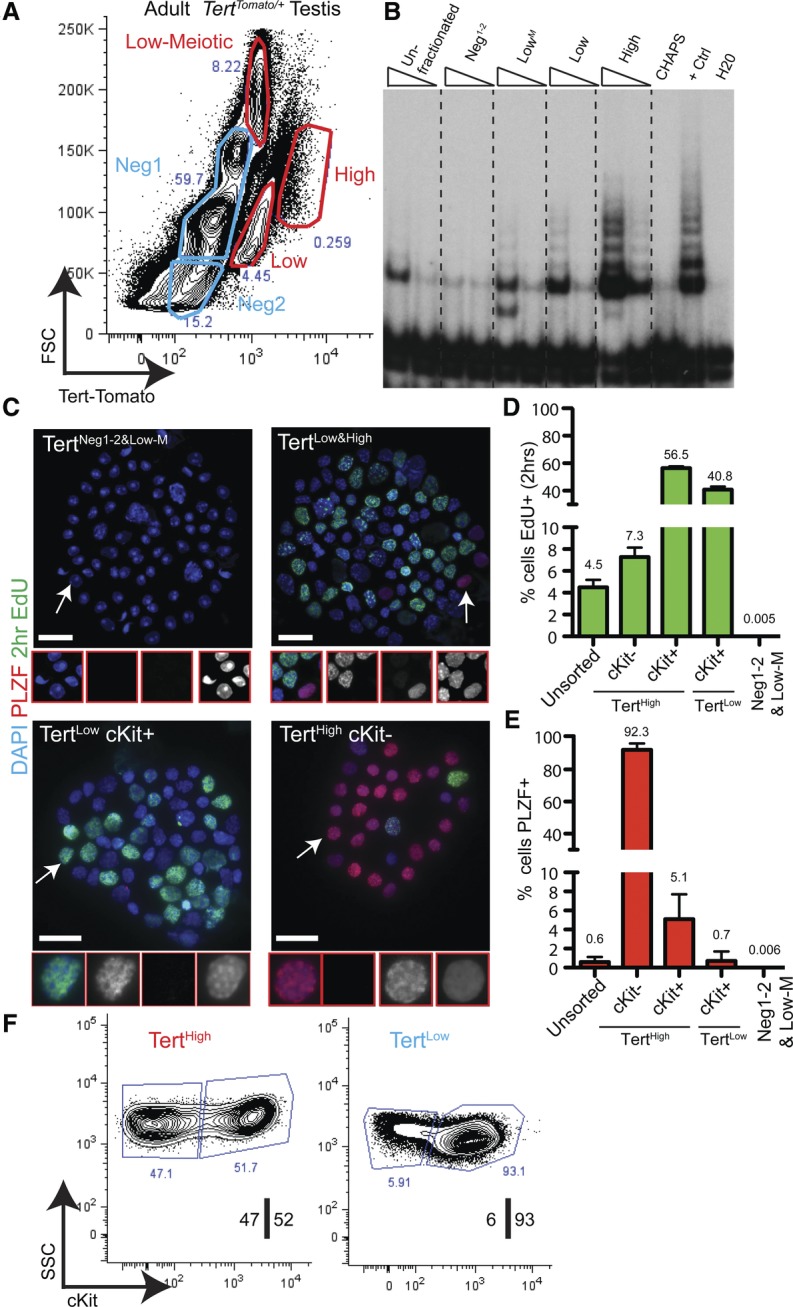
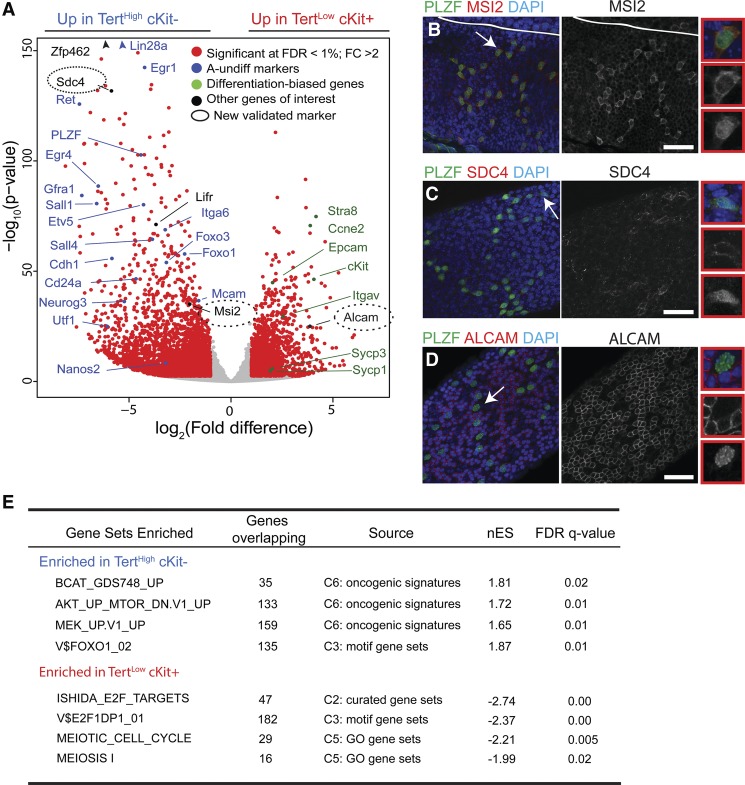
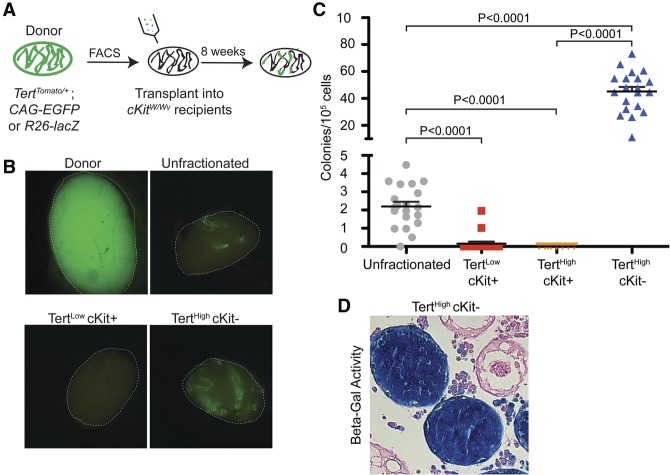
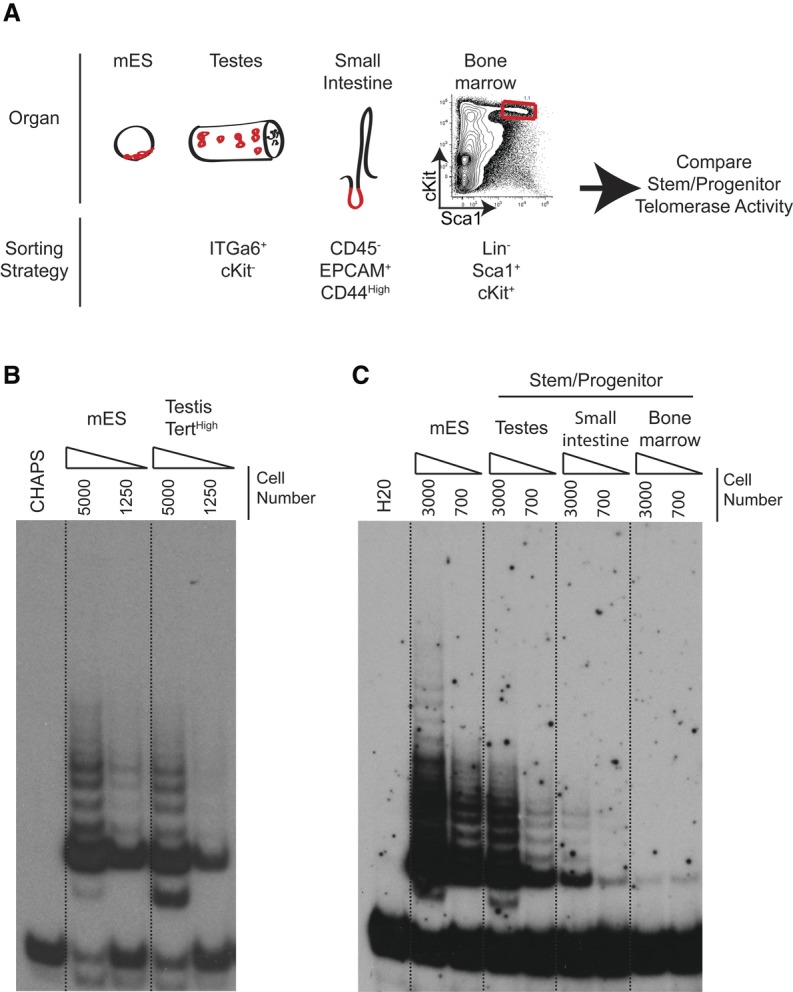
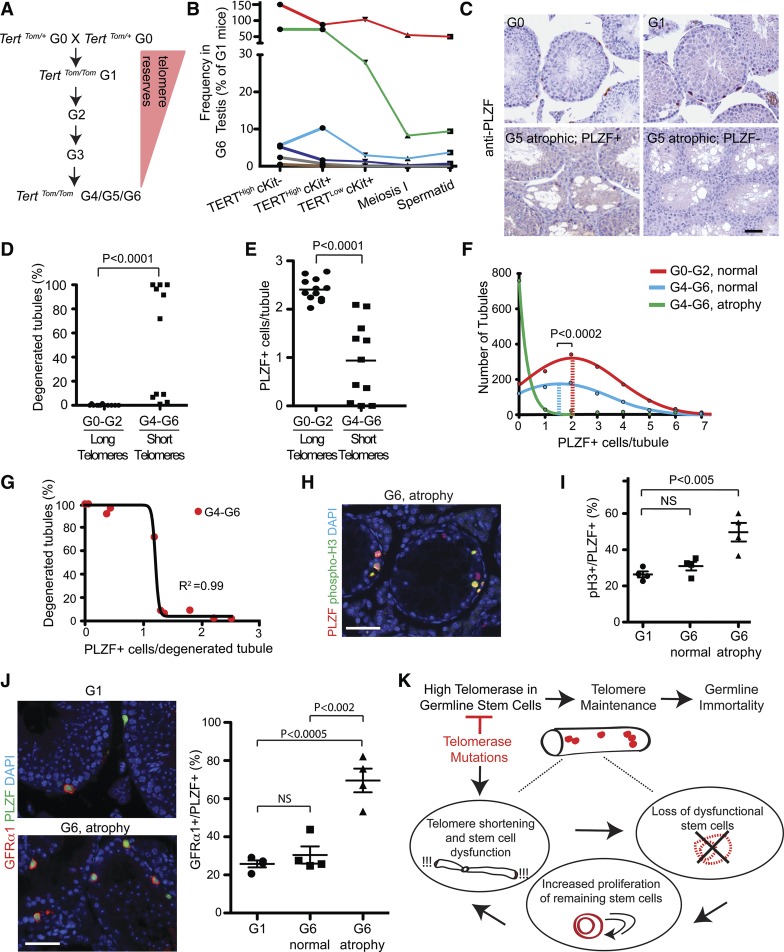
Telomerase inactivation causes loss of the male germline in worms, fish, and mice, indicating a conserved dependence on telomere maintenance in this cell lineage. Here, using telomerase reverse transcriptase (Tert) reporter mice, we found that very high telomerase expression is a hallmark of undifferentiated spermatogonia, the mitotic population where germline stem cells reside. We exploited these high telomerase levels as a basis for purifying undifferentiated spermatogonia using fluorescence-activated cell sorting. Telomerase levels in undifferentiated spermatogonia and embryonic stem cells are comparable and much greater than in somatic progenitor compartments. Within the germline, we uncovered an unanticipated gradient of telomerase activity that also enables isolation of more mature populations. Transcriptomic comparisons of Tert(High) undifferentiated spermatogonia and Tert(Low) differentiated spermatogonia by RNA sequencing reveals marked differences in cell cycle and key molecular features of each compartment. Transplantation studies show that germline stem cell activity is confined to the Tert(High) cKit(-) population. Telomere shortening in telomerase knockout strains causes depletion of undifferentiated spermatogonia and eventual loss of all germ cells after undifferentiated spermatogonia drop below a critical threshold. These data reveal that high telomerase expression is a fundamental characteristic of germline stem cells, thus explaining the broad dependence on telomerase for germline immortality in metazoans.
Keywords: aging; germline stem cells; spermatogenesis; telomerase; telomeres.
© 2015 Pech et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Similar articles
-
Clonal inactivation of TERT impairs stem cell competition.Nature. 2024 Aug;632(8023):201-208. doi: 10.1038/s41586-024-07700-w. Epub 2024 Jul 17. Nature. 2024. PMID: 39020172 Free PMC article.
-
Stage-specific embryonic antigen 4 is a membrane marker for enrichment of porcine spermatogonial stem cells.Andrology. 2020 Nov;8(6):1923-1934. doi: 10.1111/andr.12870. Epub 2020 Aug 9. Andrology. 2020. PMID: 32691968
-
Telomerase reverse transcriptase related with telomerase activity regulates tumorigenic potential of mouse embryonic stem cells.Stem Cells Dev. 2011 Jan;20(1):149-57. doi: 10.1089/scd.2009.0523. Epub 2010 Nov 5. Stem Cells Dev. 2011. PMID: 20486780
-
Telomeres in mammalian male germline cells.Int Rev Cytol. 2002;218:37-67. doi: 10.1016/s0074-7696(02)18011-9. Int Rev Cytol. 2002. PMID: 12199519 Review.
-
Regulation and Effect of Telomerase and Telomeric Length in Stem Cells.Curr Stem Cell Res Ther. 2021;16(7):809-823. doi: 10.2174/1574888X15666200422104423. Curr Stem Cell Res Ther. 2021. PMID: 32321410 Review.
Cited by
-
Unveiling the power of microenvironment in liver regeneration: an in-depth overview.Front Genet. 2023 Dec 13;14:1332190. doi: 10.3389/fgene.2023.1332190. eCollection 2023. Front Genet. 2023. PMID: 38152656 Free PMC article. Review.
-
The Transition between Telomerase and ALT Mechanisms in Hodgkin Lymphoma and Its Predictive Value in Clinical Outcomes.Cancers (Basel). 2018 May 30;10(6):169. doi: 10.3390/cancers10060169. Cancers (Basel). 2018. PMID: 29848986 Free PMC article.
-
Identification of dynamic undifferentiated cell states within the male germline.Nat Commun. 2018 Jul 19;9(1):2819. doi: 10.1038/s41467-018-04827-z. Nat Commun. 2018. PMID: 30026551 Free PMC article.
-
Gametes deficient for Pot1 telomere binding proteins alter levels of telomeric foci for multiple generations.Commun Biol. 2021 Feb 4;4(1):158. doi: 10.1038/s42003-020-01624-7. Commun Biol. 2021. PMID: 33542458 Free PMC article.
-
DNA damage in aging, the stem cell perspective.Hum Genet. 2020 Mar;139(3):309-331. doi: 10.1007/s00439-019-02047-z. Epub 2019 Jul 19. Hum Genet. 2020. PMID: 31324975 Free PMC article. Review.
References
-
- Allsopp RC, Morin GB, DePinho R, Harley CB, Weissman IL. 2003. Telomerase is required to slow telomere shortening and extend replicative lifespan of HSCs during serial transplantation. Blood 102: 517–520. - PubMed
-
- Armanios M, Chen JL, Chang YP, Brodsky RA, Hawkins A, Griffin CA, Eshleman JR, Cohen AR, Chakravarti A, Hamosh A, et al. 2005. Haploinsufficiency of telomerase reverse transcriptase leads to anticipation in autosomal dominant dyskeratosis congenita. Proc Natl Acad Sci 102: 15960–15964. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
- T32 CA009302/CA/NCI NIH HHS/United States
- NS069375/NS/NINDS NIH HHS/United States
- P01 AG036695/AG/NIA NIH HHS/United States
- R35 CA197563/CA/NCI NIH HHS/United States
- R01 CA125453/CA/NCI NIH HHS/United States
- R01 AG033747/AG/NIA NIH HHS/United States
- CA125453/CA/NCI NIH HHS/United States
- CA111691/CA/NCI NIH HHS/United States
- P30 NS069375/NS/NINDS NIH HHS/United States
- AG036695/AG/NIA NIH HHS/United States
- R01 CA111691/CA/NCI NIH HHS/United States
- AG033747/AG/NIA NIH HHS/United States
- R56 AG033747/AG/NIA NIH HHS/United States
- CA09302/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases