

Coalescence with Background and Balancing Selection in Systems with Bi- and Uniparental Reproduction: Contrasting Partial Asexuality and Selfing
- PMID: 26584901
- PMCID: PMC4701095
- DOI: 10.1534/genetics.115.181024
Coalescence with Background and Balancing Selection in Systems with Bi- and Uniparental Reproduction: Contrasting Partial Asexuality and Selfing
Abstract
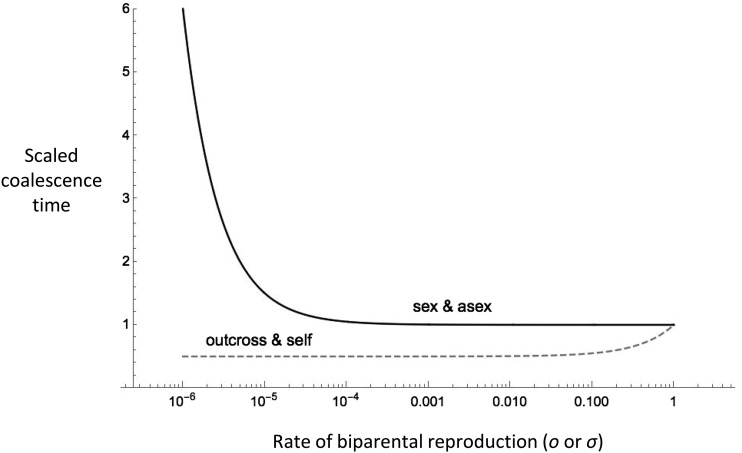
Uniparental reproduction in diploids, via asexual reproduction or selfing, reduces the independence with which separate loci are transmitted across generations. This is expected to increase the extent to which a neutral marker is affected by selection elsewhere in the genome. Such effects have previously been quantified in coalescent models involving selfing. Here we examine the effects of background selection and balancing selection in diploids capable of both sexual and asexual reproduction (i.e., partial asexuality). We find that the effect of background selection on reducing coalescent time (and effective population size) can be orders of magnitude greater when rates of sex are low than when sex is common. This is because asexuality enhances the effects of background selection through both a recombination effect and a segregation effect. We show that there are several reasons that the strength of background selection differs between systems with partial asexuality and those with comparable levels of uniparental reproduction via selfing. Expectations for reductions in Ne via background selection have been verified using stochastic simulations. In contrast to background selection, balancing selection increases the coalescence time for a linked neutral site. With partial asexuality, the effect of balancing selection is somewhat dependent upon the mode of selection (e.g., heterozygote advantage vs. negative frequency-dependent selection) in a manner that does not apply to selfing. This is because the frequency of heterozygotes, which are required for recombination onto alternative genetic backgrounds, is more dependent on the pattern of selection with partial asexuality than with selfing.
Keywords: asexual reproduction; background selection; balancing selection; coalescence; effective population size; self-fertilization.
Copyright © 2016 by the Genetics Society of America.
Figures





References
-
- Ashburner, M., 1989 Drosophila: A Laboratory Handbook. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
-
- Baer C. B., Miyamoto M. M., Denver D. R., 2007. Mutation rate variation in multicellular eukaryotes: causes and consequences. Nat. Rev. Genet. 8: 619–631. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
