

doi: 10.1016/j.antiviral.2015.11.003.
Epub 2015 Nov 14.
Teicoplanin inhibits Ebola pseudovirus infection in cell culture
Affiliations
- PMID: 26585243
- PMCID: PMC7113690
- DOI: 10.1016/j.antiviral.2015.11.003
Item in Clipboard
Teicoplanin inhibits Ebola pseudovirus infection in cell culture
Antiviral Res.
2016 Jan.
Abstract
There is currently no approved antiviral therapy for treatment of Ebola virus disease. To discover readily available approved drugs that can be rapidly repurposed for treatment of Ebola virus infections, we screened 1280 FDA-approved drugs and identified glycopeptide antibiotic teicoplanin inhibiting Ebola pseudovirus infection by blocking virus entry in the low micromolar range. Teicoplanin could be evaluated further and incorporated into ongoing clinical studies.
Keywords: Antiviral drug discovery; Ebola; High-throughput screening; Pseudovirus; Teicoplanin.
Copyright © 2015 Elsevier B.V. All rights reserved.
Figures
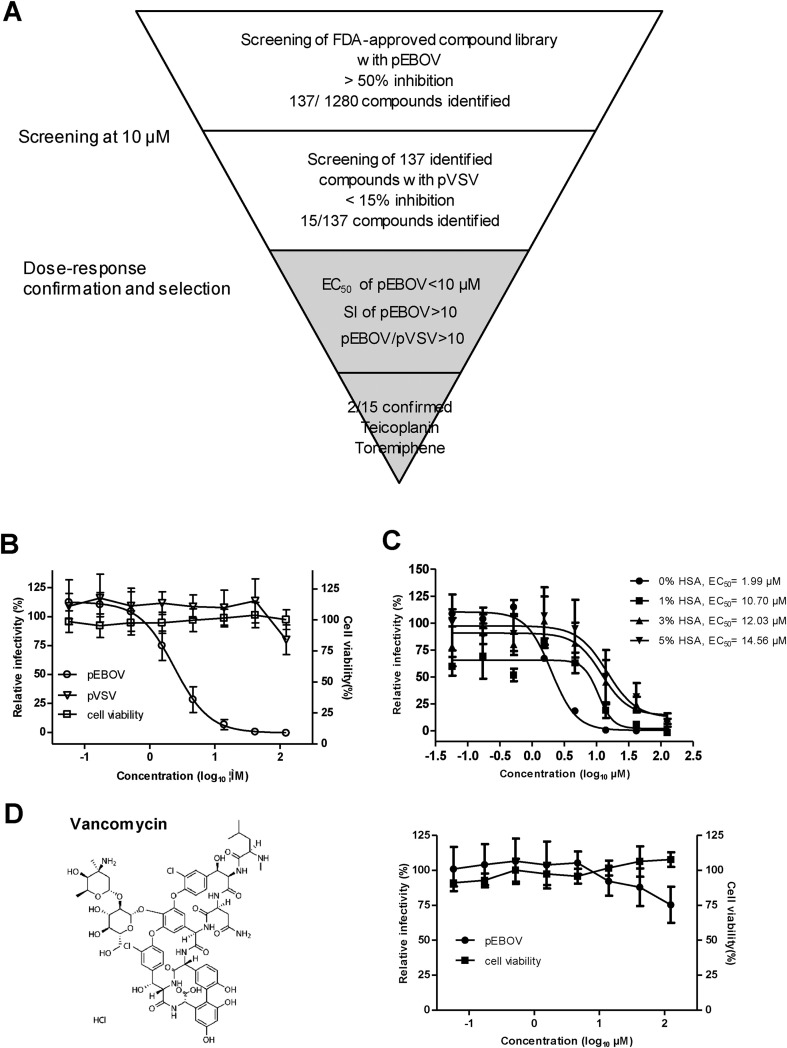
Identification of teicoplanin as an inhibitor of EBOV pseudovirus (pEBOV). (A) Flowchart of screening procedure. 1280 compounds from the FDA-approved compound library were screened in single dose at 10 μM for activity against pEBOV. 137 compounds that had activity (>50% inhibition) against pEBOV were subsequently screened against VSV pseudovirus (pVSV), leading to 15 compounds that were selectively active against pEBOV. Dose-response analysis confirmed that two compounds (teicoplanin and toremiphene) met the selection criteria of EC50 < 10 μM, SI > 10 against pEBOV, and pEBOV/pVSV > 10. (B) Activity of teicoplanin against pEBOV and pVSV. Three-fold serial dilutions of teicoplanin were added to Vero cells, after 72 h of incubation, the relative infectivities were analyzed by measuring the luciferase and presented as a percentage of luciferase derived from the compound-treated cells compared with that from the mock-treated cells. Cytotoxicity was also examined by incubation of Vero cells with the indicated concentrations of teicoplanin and was presented as a percentage of luminescence derived from the compound-treated cells compared with that from the mock-treated cells (with medium). (C) Effect of human serum albumin (HSA) on the anti-pEBOV activity of teicoplanin. Activity of teicoplanin against pEBOV was evaluated in the presence of indicated concentrations of HSA and EC50s were calculated using Prism's nonlinear regression (GraphPadPrism5). (D) Activity of vancomycin against pEBOV. Left, chemical structure of vancomycin; right, activity of vancomycin against pEBOV and cytotoxicity of vancomycin. For (B), (C), and (D), average results from three experiments are shown. Error bars represent the standard deviations (B and D) or standard error (C) of means of three independent measurements.
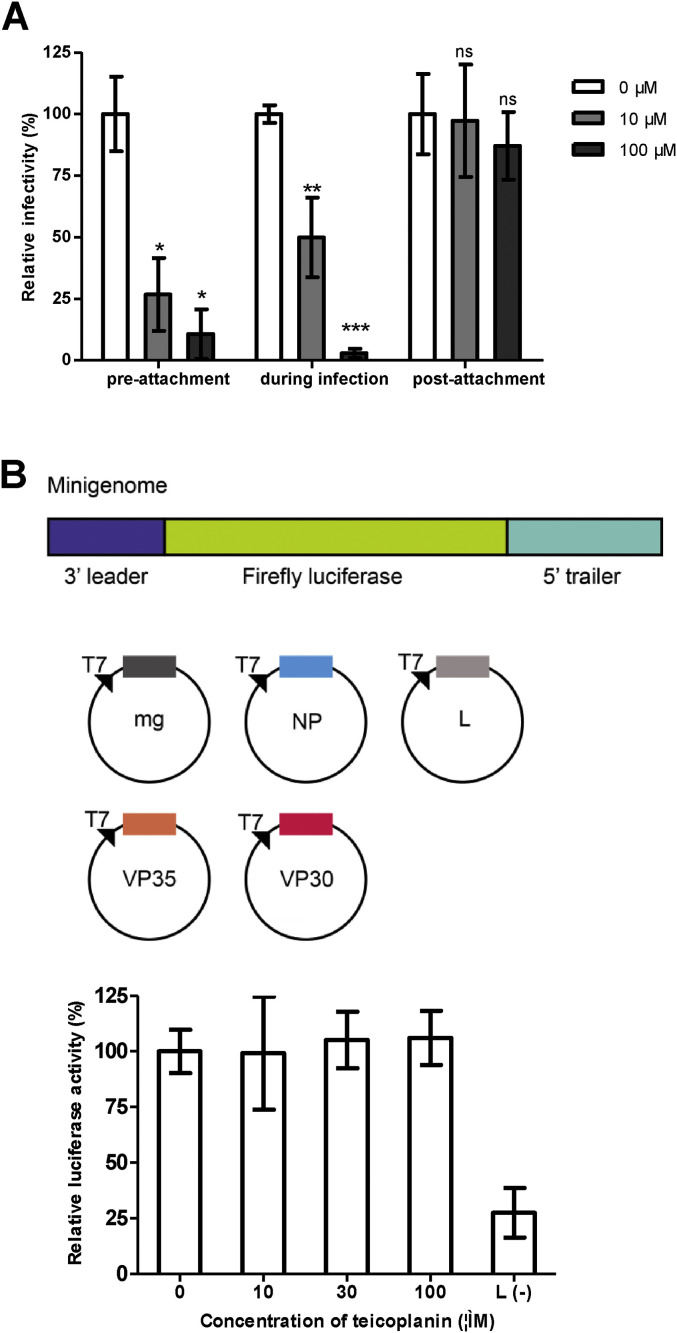
Teicoplanin blocks pEBOV entry but does not inhibit viral RNA replication. (A) Time-of-addition studies with teicoplanin. Vero cells were infected with pEBOV at 37 °C for 1 h in the presence (during infection) of 10 or 100 μM of teicoplanin and washed to remove unbound pEBOV and compounds, and then fresh medium was added and incubated at 37 °C for 72 h. For the pre-attachment assay, pEBOV was incubated with 10 or 100 μM of teicoplanin at 4 °C for 1 h prior to addition to cells. The virus-compound mixture was incubated with cells at 37 °C for 1 h, after which the cells were washed with medium and incubated at 37 °C for 72 h. For the post-attachment assay, Vero cells were prechilled to 4 °C, and pEBOV was added to cells, and virus adsorption was allowed for 1 h at 4 °C. Cells were washed three times with cold medium to remove unbound virus, and 10 or 100 μM of teicoplanin were added for 1 h at 4 °C, and cells then were washed and incubated at 37 °C for 72 h. The data are the averages for quadruplicate wells for two independent experiments, and the error bars represent the standard deviations of the means. Statistical significance between treated and control group was analyzed by t-test (ns, not significant, *, p < 0.05, **, p < 0.01 ***, p < 0.001). (B) Minigenome assay with teicoplanin. Upper, schematic diagram of the Ebola virus (Zaire strain) minigenome used in this study, in which the viral genes are removed and replaced with a firefly luciferase (FLuc) reporter gene, but the nontranscribed leader and trailer regions as well as the noncoding regions upstream and downstream of NP gene are retained. The minigenome (mg) was flanked by the T7 RNA polymerase promoter (T7) and a ribozyme (HDVr). The minigenome is replicated and transcribed by NP, VP35, VP30, and L provided in trans from expression plasmids. Lower, BSR T7/5 cells were transfected with minigenome and expression plasmids for the RNP proteins NP, VP35, VP30, and L, and after 2 h of incubation with transfection complex, cells were treated with 10 μM, 30 μM, or 100 μM of teicoplanin. Firefly and renilla luciferase activities were measured at 24 h post-transfection. Cells transfected without L plasmid were set as background. The effects of teicoplanin were presented as a percentage of firefly luciferase (normalized against renilla luciferase) derived from the compound-treated cells compared with that from the mock-treated cells. The data presented were obtained from two independent experiments. Error bars represent the standard deviations from two independent experiments.
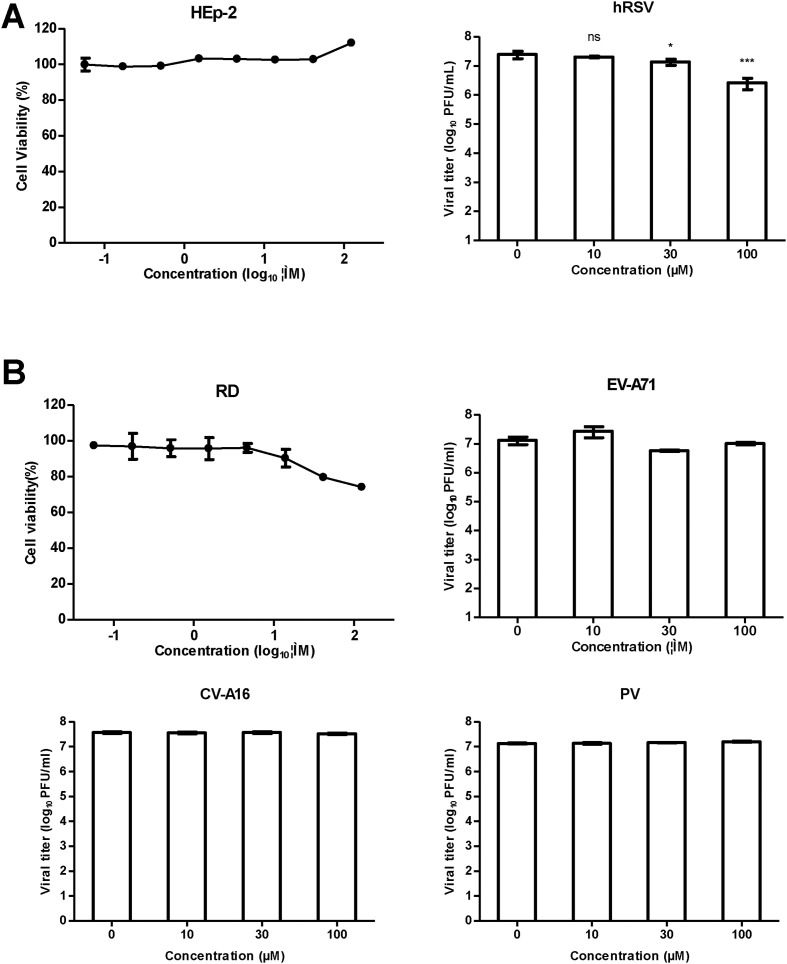
Effect of teicoplanin on hRSV, EV-A71, CV-A16, and PV1. (A) Left, cytotoxicity of teicoplanin on HEp-2 cells was assessed in eight-point dose-response. Right, HEp-2 cells were infected with hRSV (subtype A, strain Long) at an MOI of 0.1 and treated with teicoplanin at the indicated concentrations. Supernatants were collected at 48 h post-infection, and viral titers were determined by immunostaining assay. (B) Cytotoxicity of teicoplanin on RD cells was assessed in eight-point dose-response. Cells were infected with EV-A71 (MOI = 0.1), CV-A16, and PV1 (MOI = 0.01) and treated with teicoplanin at the indicated concentrations. Supernatants were collected at 42 h post-infection, and viral titers were determined by plaque assay. For (A) and (B), the data presented were obtained from two independent experiments. Error bars represent the standard deviations from two independent experiments. Statistical significance between treated and control group was analyzed by t-test (ns, not significant, *, p < 0.05, ***, p < 0.001).
References
-
- Altmeyer R. Virus attachment and entry offer numerous targets for antiviral therapy. Curr. Pharm. Des. 2004;10:3701–3712. - PubMed
-
- Ashburn T.T., Thor K.B. Drug repositioning: identifying and developing new uses for existing drugs. Nature reviews. Drug Discov. 2004;3:673–683. - PubMed
-
- Baize S., Pannetier D., Oestereich L., Rieger T., Koivogui L., Magassouba N., Soropogui B., Sow M.S., Keita S., De Clerck H., Tiffany A., Dominguez G., Loua M., Traore A., Kolie M., Malano E.R., Heleze E., Bocquin A., Mely S., Raoul H., Caro V., Cadar D., Gabriel M., Pahlmann M., Tappe D., Schmidt-Chanasit J., Impouma B., Diallo A.K., Formenty P., Van Herp M., Gunther S. Emergence of Zaire Ebola virus disease in Guinea. N. Engl. J. Med. 2014;371:1418–1425. - PubMed
-
- Balzarini J., Keyaerts E., Vijgen L., Egberink H., De Clercq E., Van Ranst M., Printsevskaya S.S., Olsufyeva E.N., Solovieva S.E., Preobrazhenskaya M.N. Inhibition of feline (FIPV) and human (SARS) coronavirus by semisynthetic derivatives of glycopeptide antibiotics. Antivir. Res. 2006;72:20–33. - PMC - PubMed
-
- Balzarini J., Pannecouque C., De Clercq E., Pavlov A.Y., Printsevskaya S.S., Miroshnikova O.V., Reznikova M.I., Preobrazhenskaya M.N. Antiretroviral activity of semisynthetic derivatives of glycopeptide antibiotics. J. Med. Chem. 2003;46:2755–2764. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
