

The endocannabinoid system in guarding against fear, anxiety and stress
- PMID: 26585799
- PMCID: PMC5871913
- DOI: 10.1038/nrn4036
The endocannabinoid system in guarding against fear, anxiety and stress
Abstract
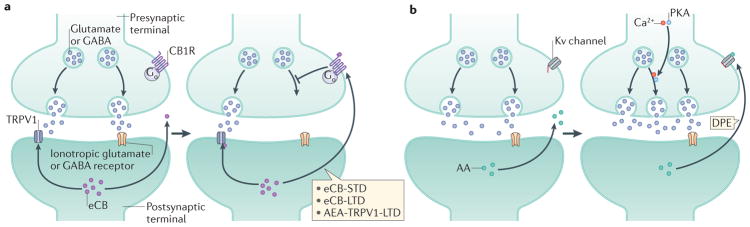
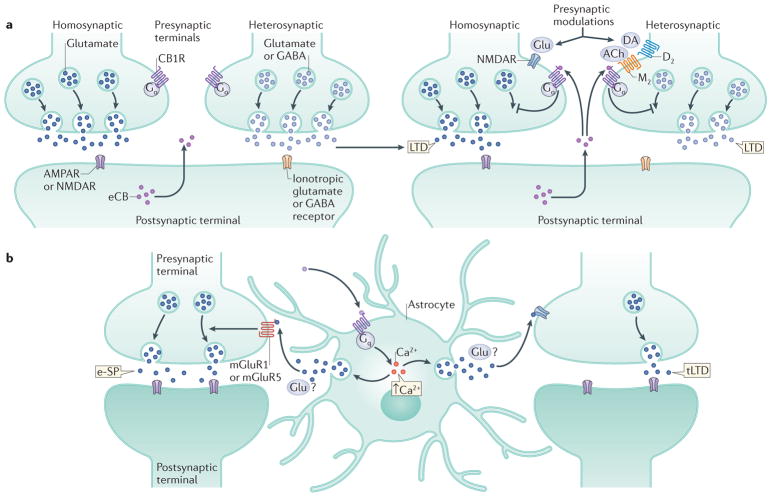
The endocannabinoid (eCB) system has emerged as a central integrator linking the perception of external and internal stimuli to distinct neurophysiological and behavioural outcomes (such as fear reaction, anxiety and stress-coping), thus allowing an organism to adapt to its changing environment. eCB signalling seems to determine the value of fear-evoking stimuli and to tune appropriate behavioural responses, which are essential for the organism's long-term viability, homeostasis and stress resilience; and dysregulation of eCB signalling can lead to psychiatric disorders. An understanding of the underlying neural cell populations and cellular processes enables the development of therapeutic strategies to mitigate behavioural maladaptation.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Tart CT. Marijuana intoxication common experiences. Nature. 1970;226:701–704. - PubMed
-
- Lafenetre P, Chaouloff F, Marsicano G. The endocannabinoid system in the processing of anxiety and fear and how CB1 receptors may modulate fear extinction. Pharmacol Res. 2007;56:367–381. - PubMed
-
- Riebe CJ, Pamplona FA, Kamprath K, Wotjak CT. Fear relief—toward a new conceptual frame work and what endocannabinoids gotta do with it. Neuroscience. 2012;204:159–185. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
