

Convergence, Divergence, and Reconvergence in a Feedforward Network Improves Neural Speed and Accuracy
- PMID: 26586183
- PMCID: PMC5488793
- DOI: 10.1016/j.neuron.2015.10.018
Convergence, Divergence, and Reconvergence in a Feedforward Network Improves Neural Speed and Accuracy
Abstract
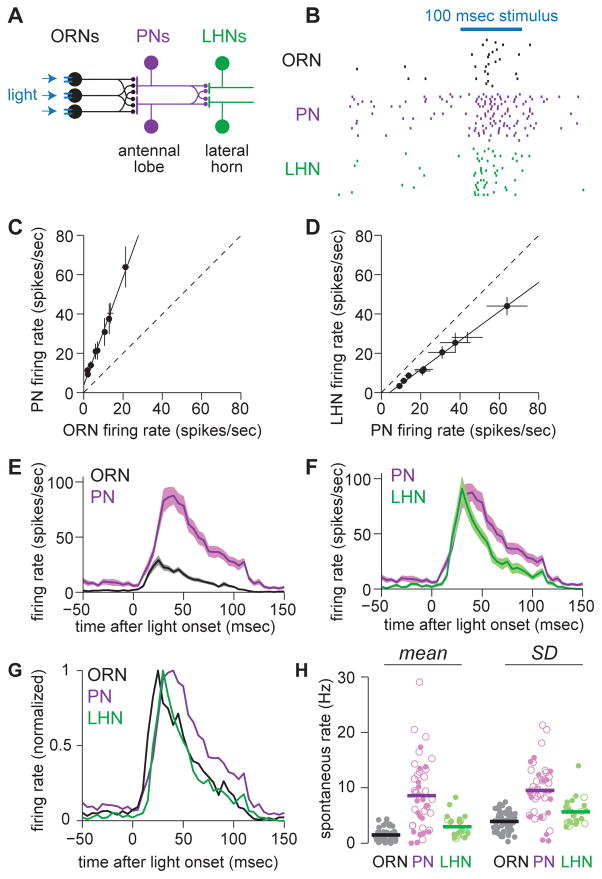
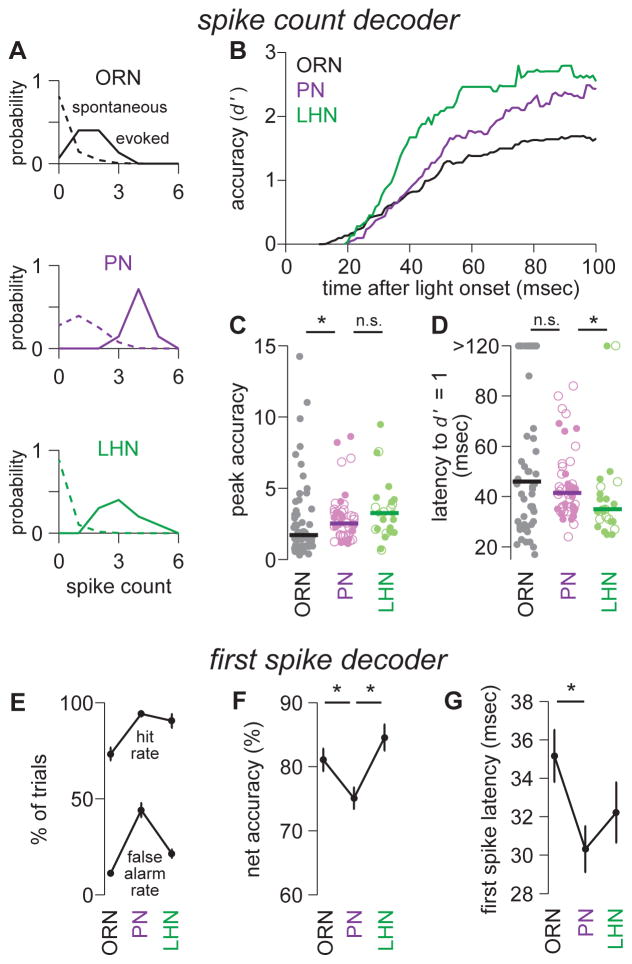
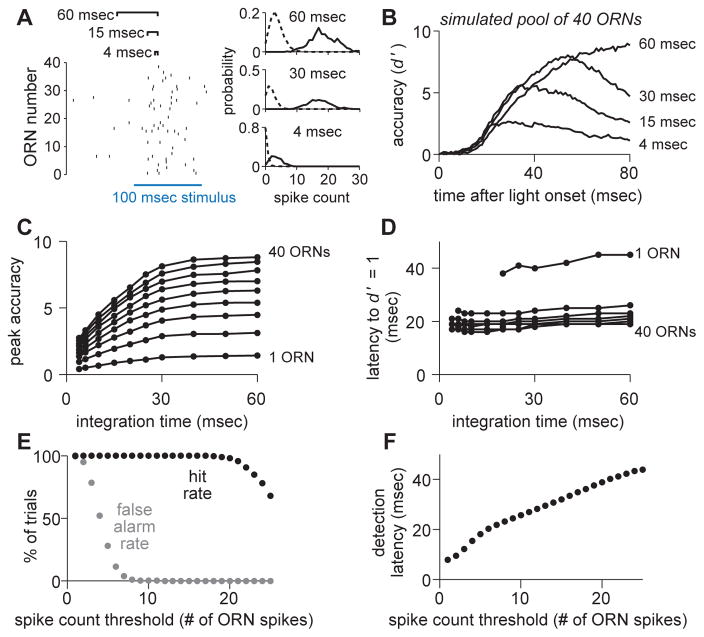
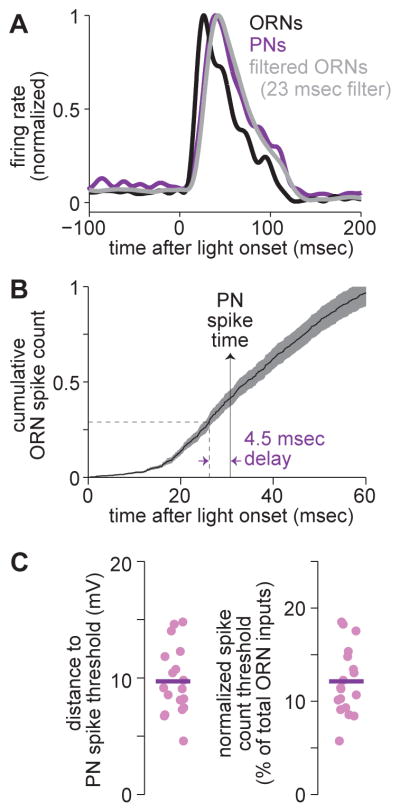
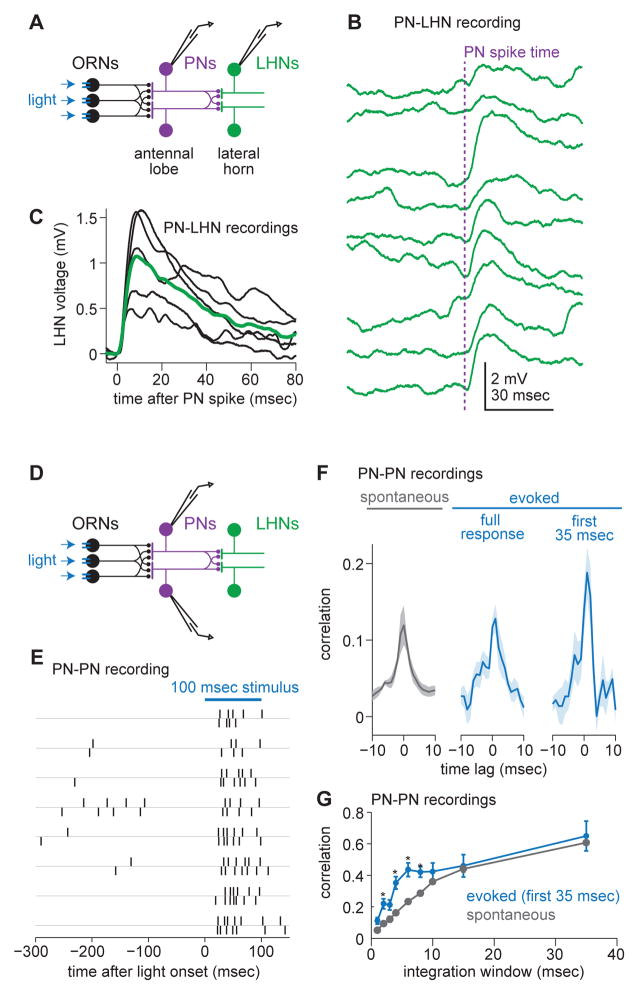
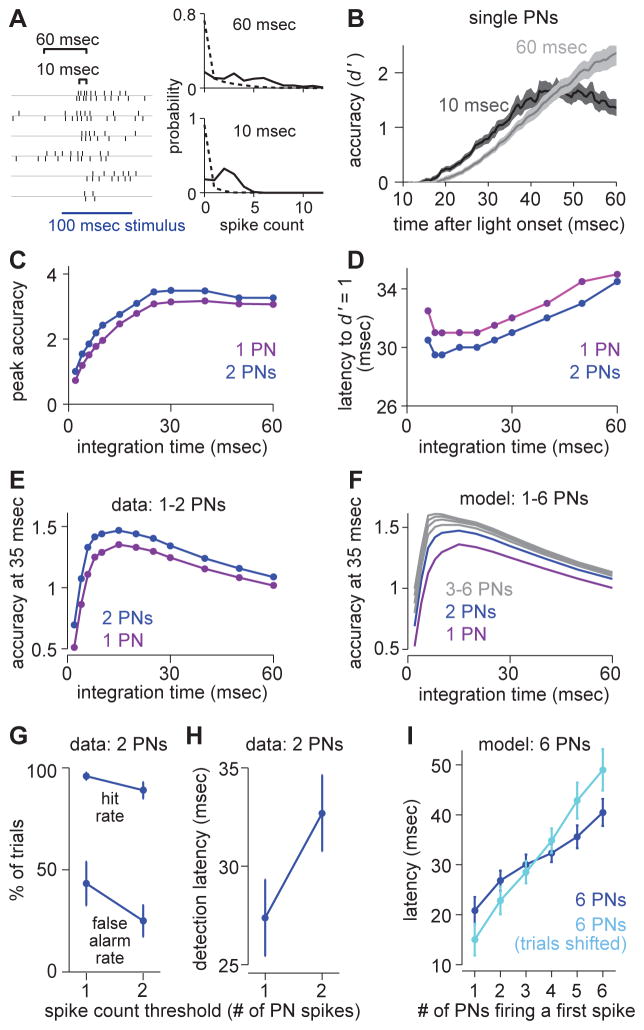
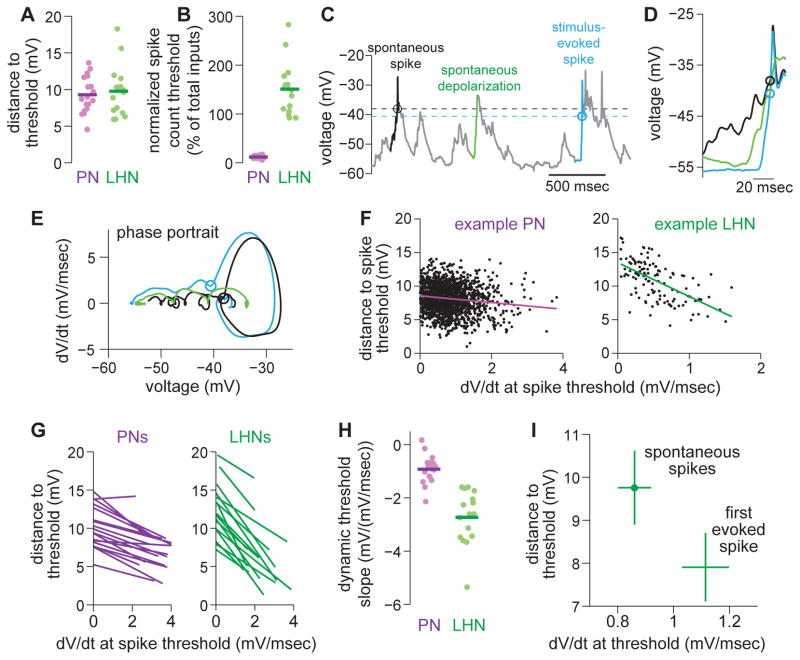
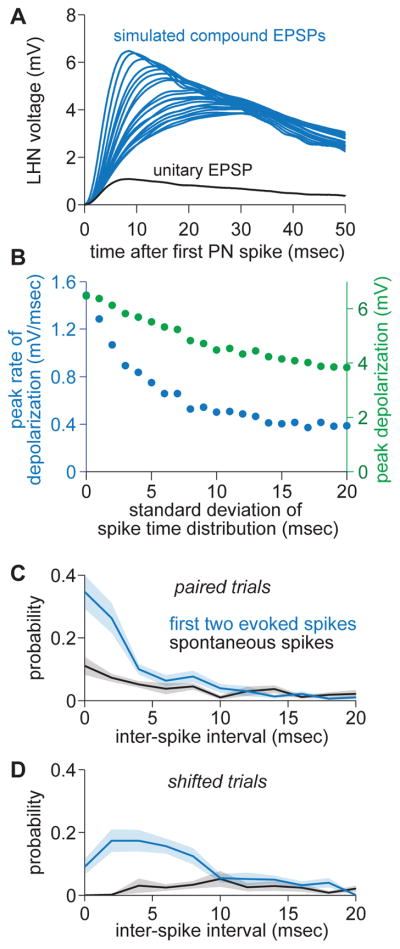
One of the proposed canonical circuit motifs employed by the brain is a feedforward network where parallel signals converge, diverge, and reconverge. Here we investigate a network with this architecture in the Drosophila olfactory system. We focus on a glomerulus whose receptor neurons converge in an all-to-all manner onto six projection neurons that then reconverge onto higher-order neurons. We find that both convergence and reconvergence improve the ability of a decoder to detect a stimulus based on a single neuron's spike train. The first transformation implements averaging, and it improves peak detection accuracy but not speed; the second transformation implements coincidence detection, and it improves speed but not peak accuracy. In each case, the integration time and threshold of the postsynaptic cell are matched to the statistics of convergent spike trains.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
Comment in
-
I Want It All and I Want It Now: How a Neural Circuit Encodes Odor with Speed and Accuracy.Neuron. 2015 Dec 2;88(5):852-854. doi: 10.1016/j.neuron.2015.11.016. Neuron. 2015. PMID: 26637793
References
-
- Alonso JM, Usrey WM, Reid RC. Precisely correlated firing in cells of the lateral geniculate nucleus. Nature. 1996;383:815–819. - PubMed
-
- Averbeck BB, Latham PE, Pouget A. Neural correlations, population coding and computation. Nat Rev Neurosci. 2006;7:358–366. - PubMed
-
- Bargmann CI. Comparative chemosensation from receptors to ecology. Nature. 2006;444:295–301. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
