

Structural basis for leucine sensing by the Sestrin2-mTORC1 pathway
- PMID: 26586190
- PMCID: PMC4698039
- DOI: 10.1126/science.aad2087
Structural basis for leucine sensing by the Sestrin2-mTORC1 pathway
Abstract
Eukaryotic cells coordinate growth with the availability of nutrients through the mechanistic target of rapamycin complex 1 (mTORC1), a master growth regulator. Leucine is of particular importance and activates mTORC1 via the Rag guanosine triphosphatases and their regulators GATOR1 and GATOR2. Sestrin2 interacts with GATOR2 and is a leucine sensor. Here we present the 2.7 angstrom crystal structure of Sestrin2 in complex with leucine. Leucine binds through a single pocket that coordinates its charged functional groups and confers specificity for the hydrophobic side chain. A loop encloses leucine and forms a lid-latch mechanism required for binding. A structure-guided mutation in Sestrin2 that decreases its affinity for leucine leads to a concomitant increase in the leucine concentration required for mTORC1 activation in cells. These results provide a structural mechanism of amino acid sensing by the mTORC1 pathway.
Copyright © 2016, American Association for the Advancement of Science.
Figures
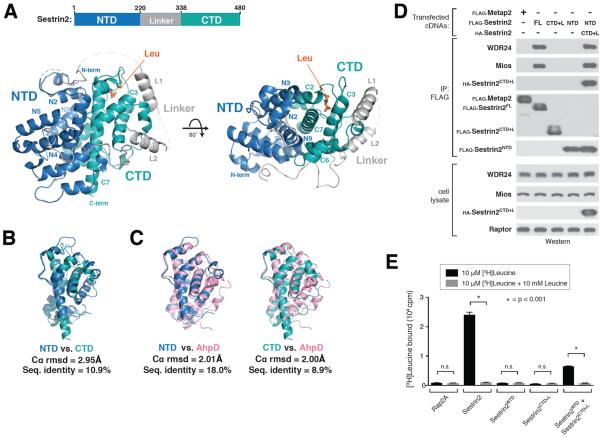
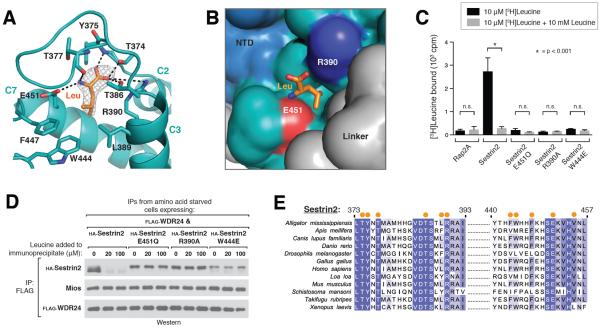
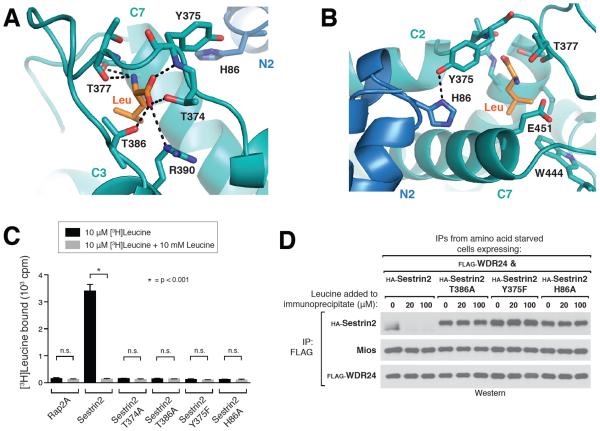
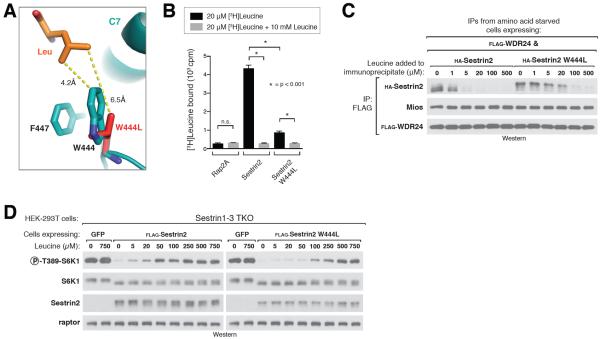
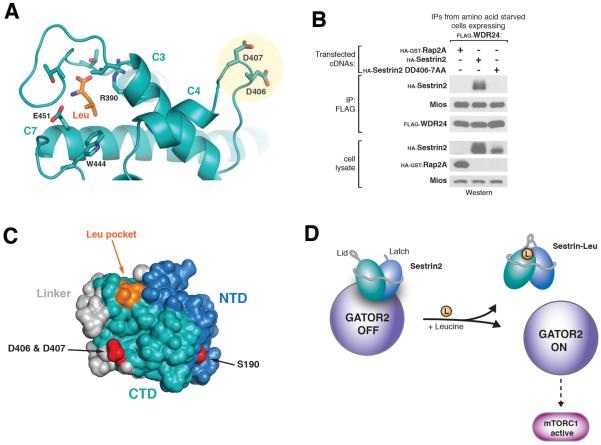
Comment in
-
CELL SIGNALING. Seeing mTORC1 specificity.Science. 2016 Jan 1;351(6268):25-6. doi: 10.1126/science.aad9696. Science. 2016. PMID: 26721988 No abstract available.
References
-
- Potier M, Darcel N, Tomé D. Protein, amino acids and the control of food intake. Current Opinion in Clinical Nutrition and Metabolic Care. 2009;12:54–58. - PubMed
-
- Greiwe JS, Kwon G, McDaniel ML, Semenkovich CF. Leucine and insulin activate p70 S6 kinase through different pathways in human skeletal muscle. American journal of physiology. Endocrinology and metabolism. 2001;281:E466–471. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- R01 AI047389/AI/NIAID NIH HHS/United States
- R01 CA103866/CA/NCI NIH HHS/United States
- T32 GM007287/GM/NIGMS NIH HHS/United States
- P41 GM103403/GM/NIGMS NIH HHS/United States
- F30 CA189333/CA/NCI NIH HHS/United States
- P30 CA014051/CA/NCI NIH HHS/United States
- T32 GM007753/GM/NIGMS NIH HHS/United States
- F31 CA180271/CA/NCI NIH HHS/United States
- T32GM007287/GM/NIGMS NIH HHS/United States
- R37 AI047389/AI/NIAID NIH HHS/United States
- R01CA103866/CA/NCI NIH HHS/United States
- Howard Hughes Medical Institute/United States
- F31 CA189437/CA/NCI NIH HHS/United States
- S10 RR029205/RR/NCRR NIH HHS/United States
- AI47389/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
