

Selection and dynamics of embryonic stem cell integration into early mouse embryos
- PMID: 26586221
- PMCID: PMC4725202
- DOI: 10.1242/dev.124602
Selection and dynamics of embryonic stem cell integration into early mouse embryos
Abstract
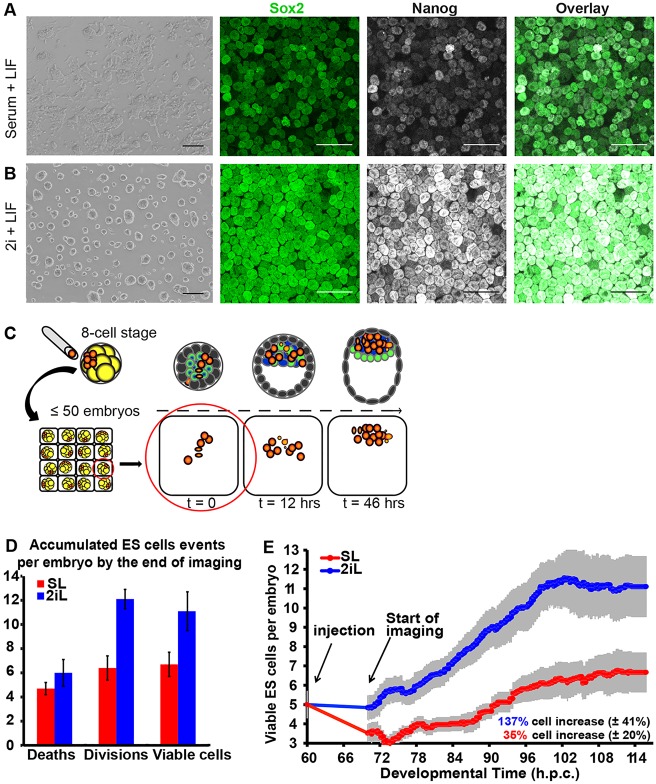
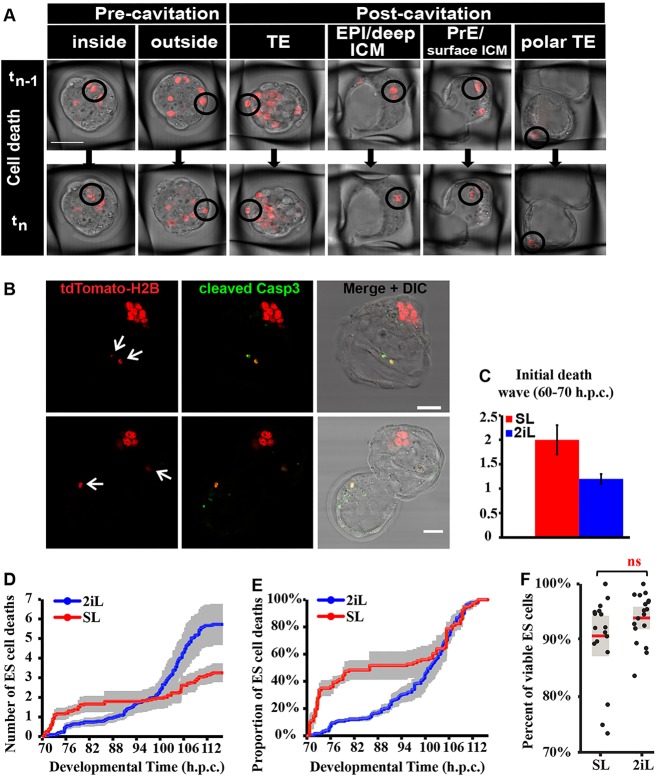
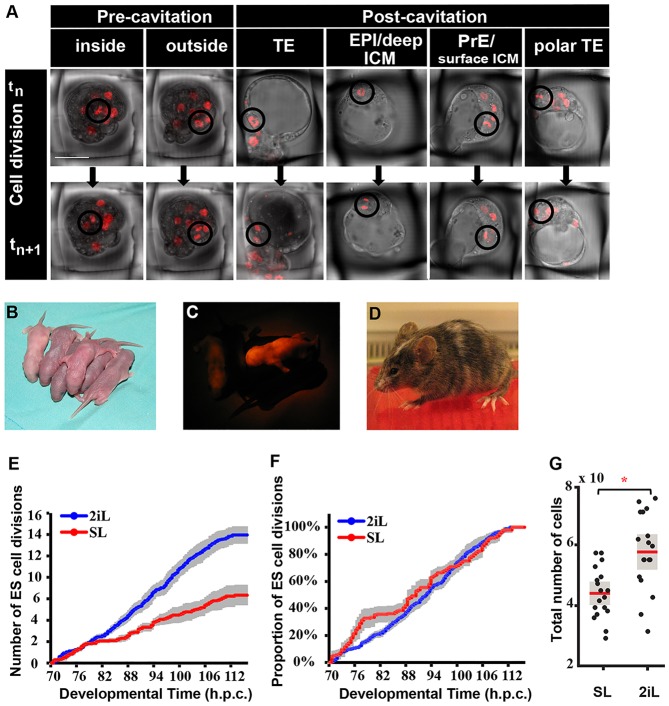
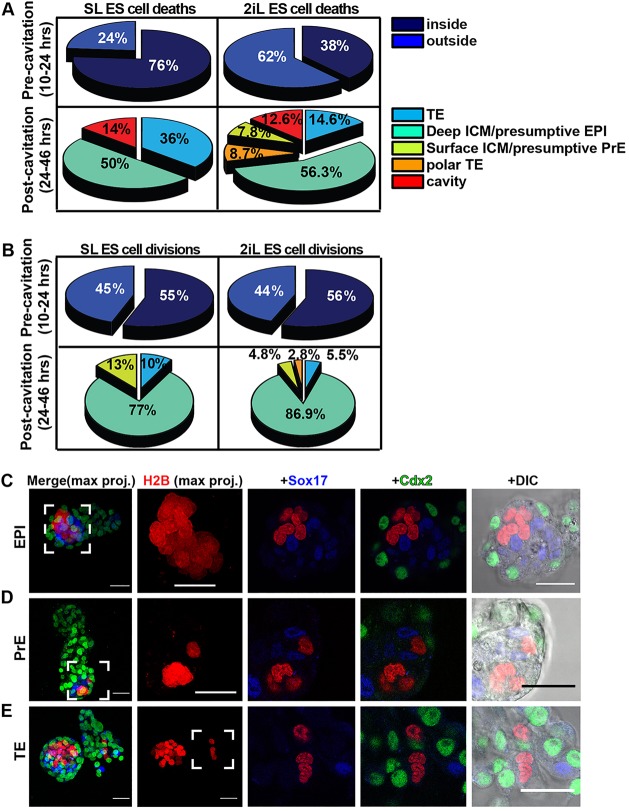
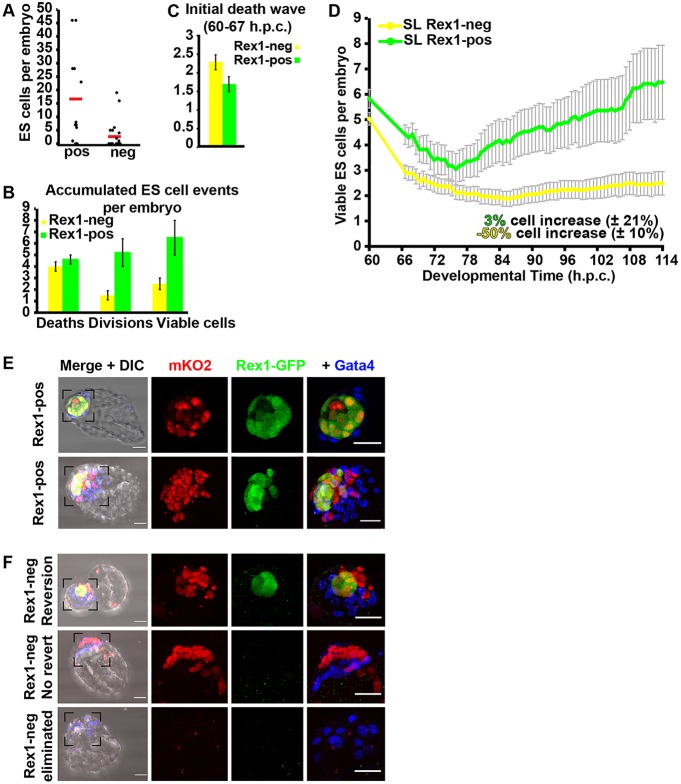
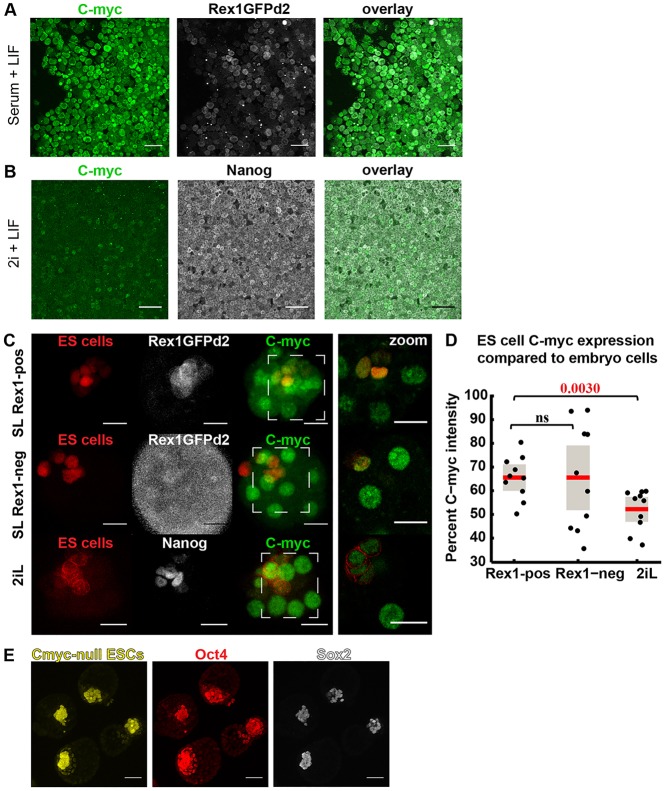
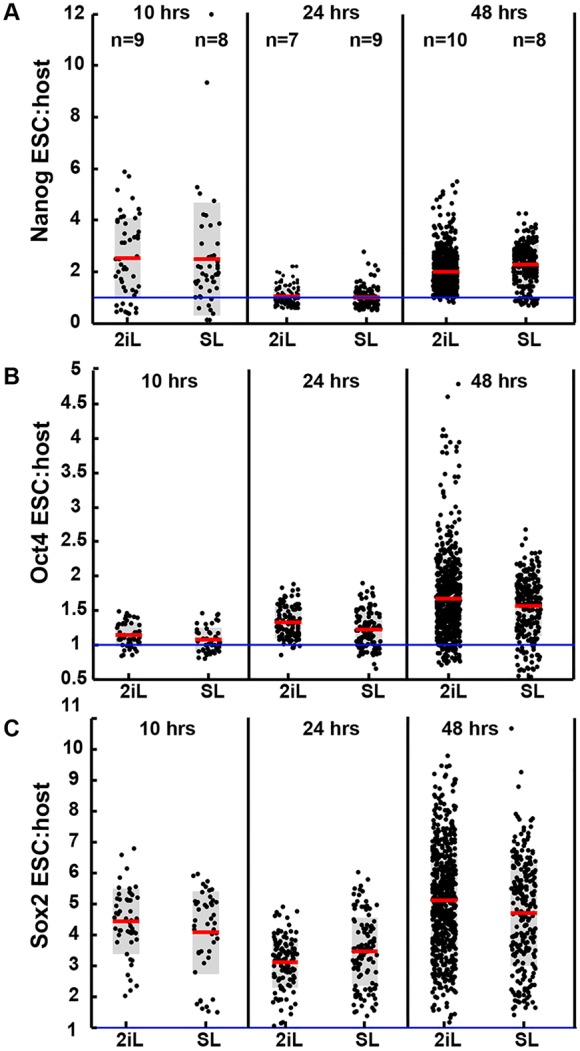
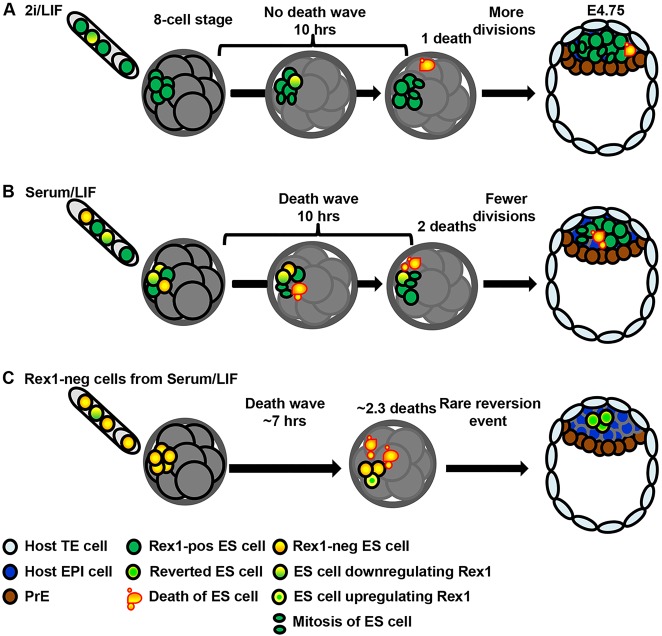
The process by which pluripotent cells incorporate into host embryos is of interest to investigate cell potency and cell fate decisions. Previous studies suggest that only a minority of the embryonic stem cell (ESC) inoculum contributes to the adult chimaera. How incoming cells are chosen for integration or elimination remains unclear. By comparing a heterogeneous mix of undifferentiated and differentiating ESCs (serum/LIF) with more homogeneous undifferentiated culture (2i/LIF), we examine the role of cellular heterogeneity in this process. Time-lapse ex vivo imaging revealed a drastic elimination of serum/LIF ESCs during early development in comparison with 2i/LIF ESCs. Using a fluorescent reporter for naive pluripotency (Rex1-GFP), we established that the acutely eliminated serum/LIF ESCs had started to differentiate. The rejected cells were apparently killed by apoptosis. We conclude that a selection process exists by which unwanted differentiating cells are eliminated from the embryo. However, occasional Rex1(-) cells were able to integrate. Upregulation of Rex1 occurred in a proportion of these cells, reflecting the potential of the embryonic environment to expedite diversion from differentiation priming to enhance the developing embryonic epiblast.
Keywords: Chimaera; Embryonic stem cell; Live imaging; Mouse blastocyst; Pluripotency.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
-
- Beddington R. S. and Robertson E. J. (1989). An assessment of the developmental potential of embryonic stem cells in the midgestation mouse embryo. Development 105, 733-737. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
