

Dendritic Cell-Specific Deletion of β-Catenin Results in Fewer Regulatory T-Cells without Exacerbating Autoimmune Collagen-Induced Arthritis
- PMID: 26587585
- PMCID: PMC4654567
- DOI: 10.1371/journal.pone.0142972
Dendritic Cell-Specific Deletion of β-Catenin Results in Fewer Regulatory T-Cells without Exacerbating Autoimmune Collagen-Induced Arthritis
Abstract
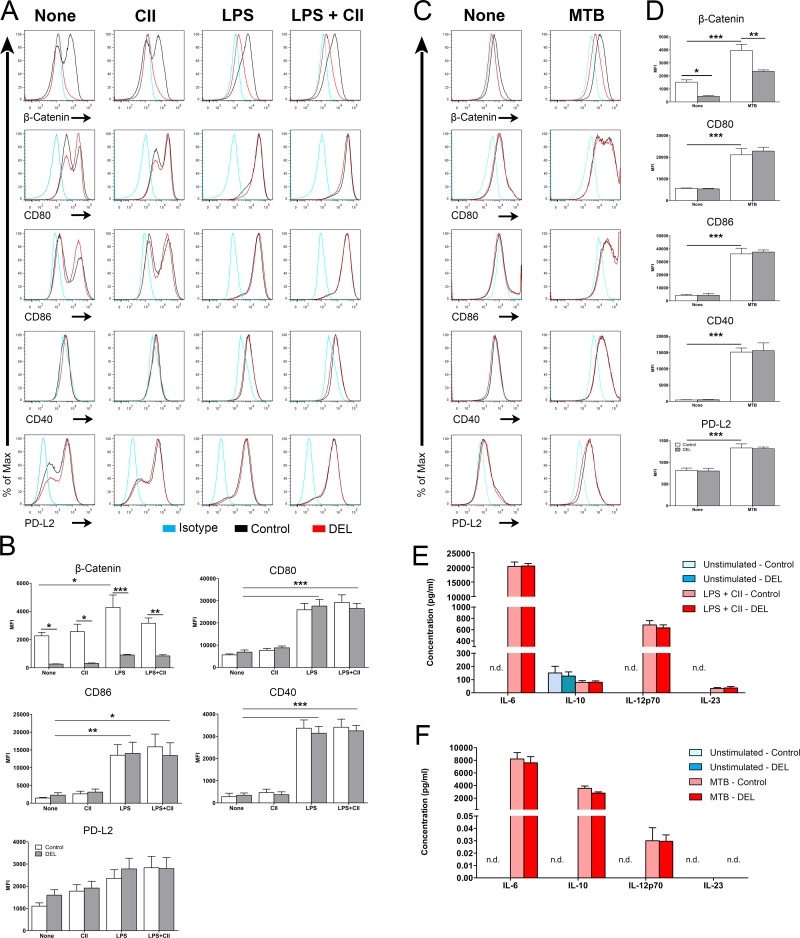
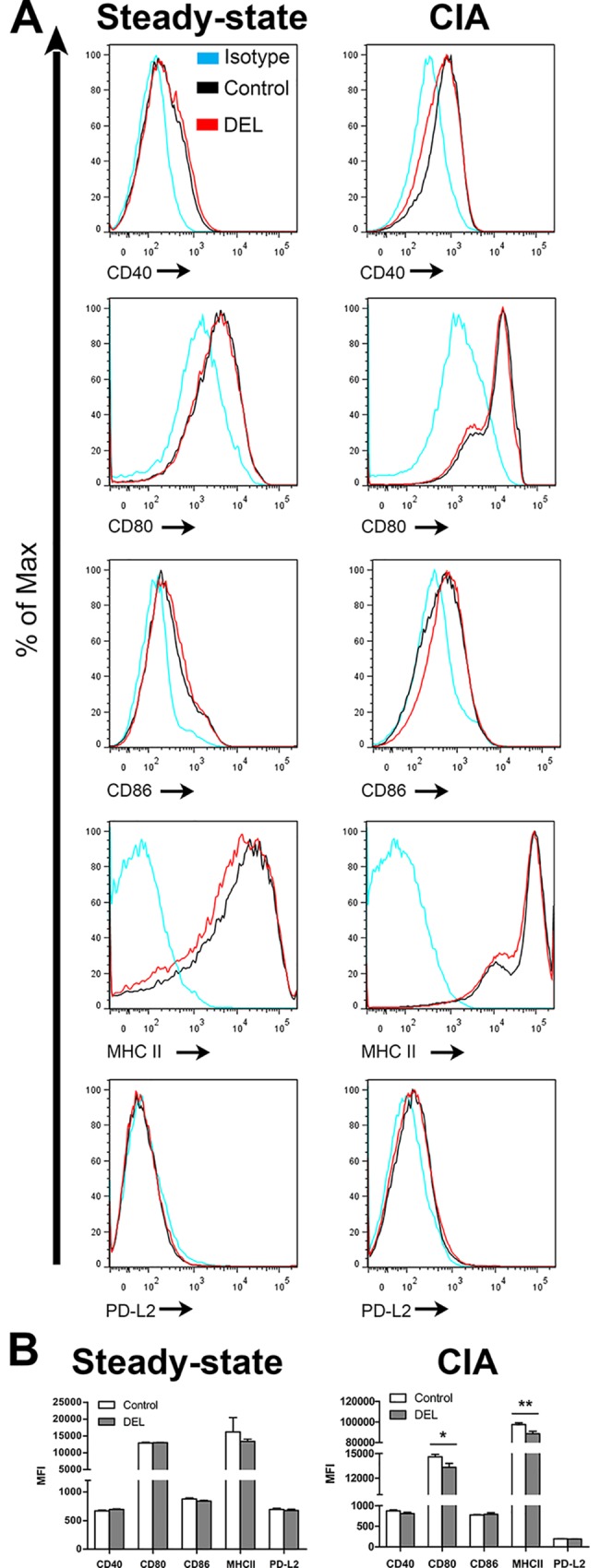
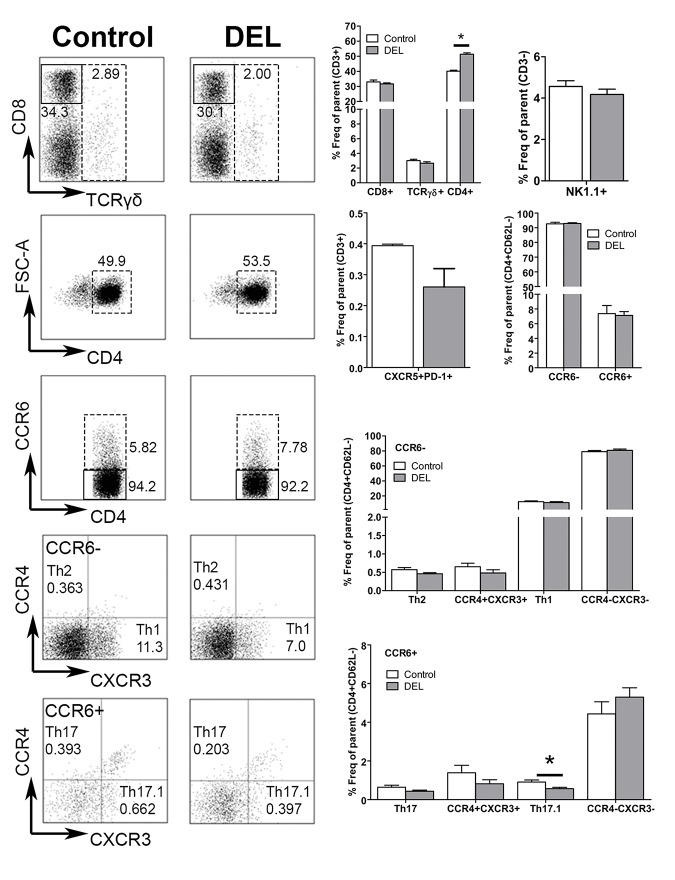
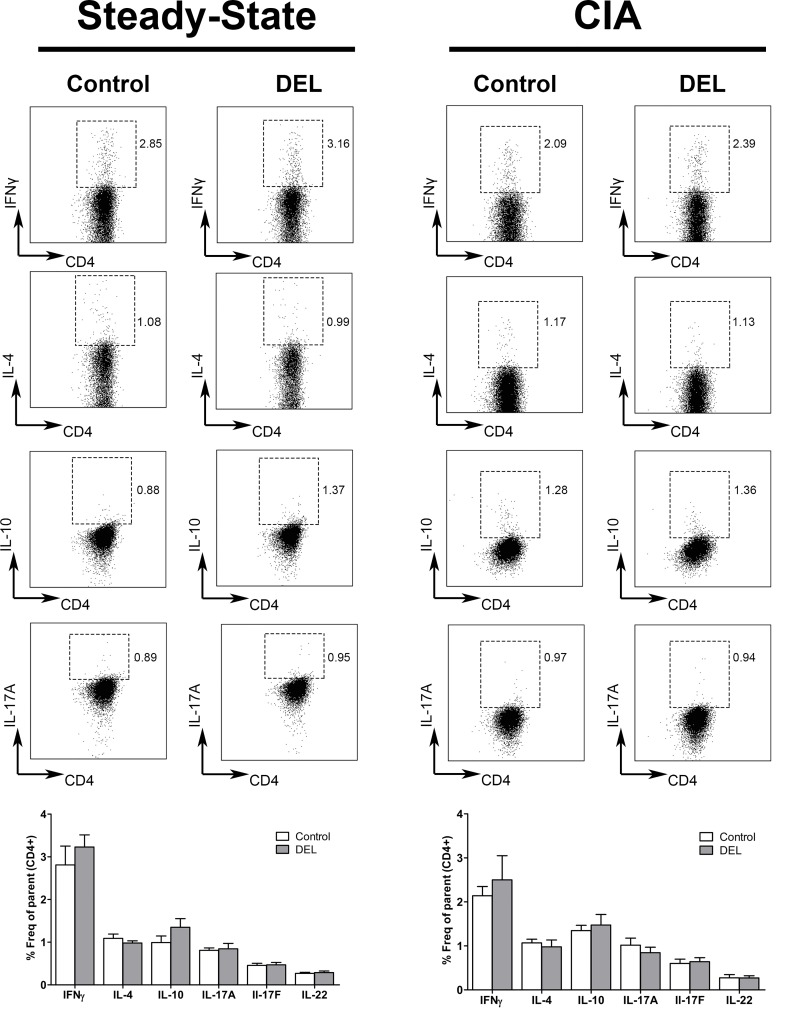
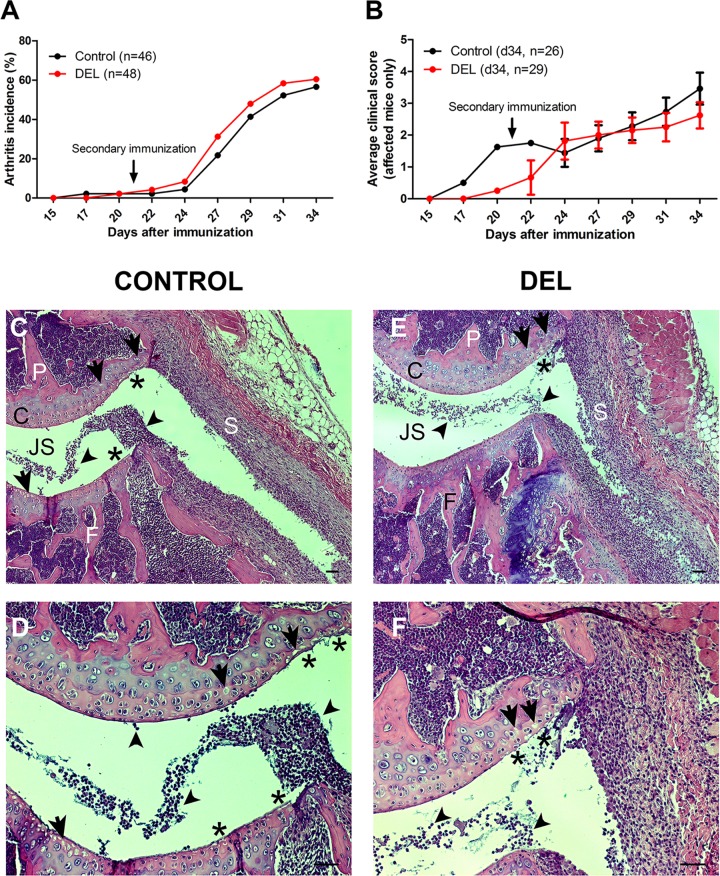
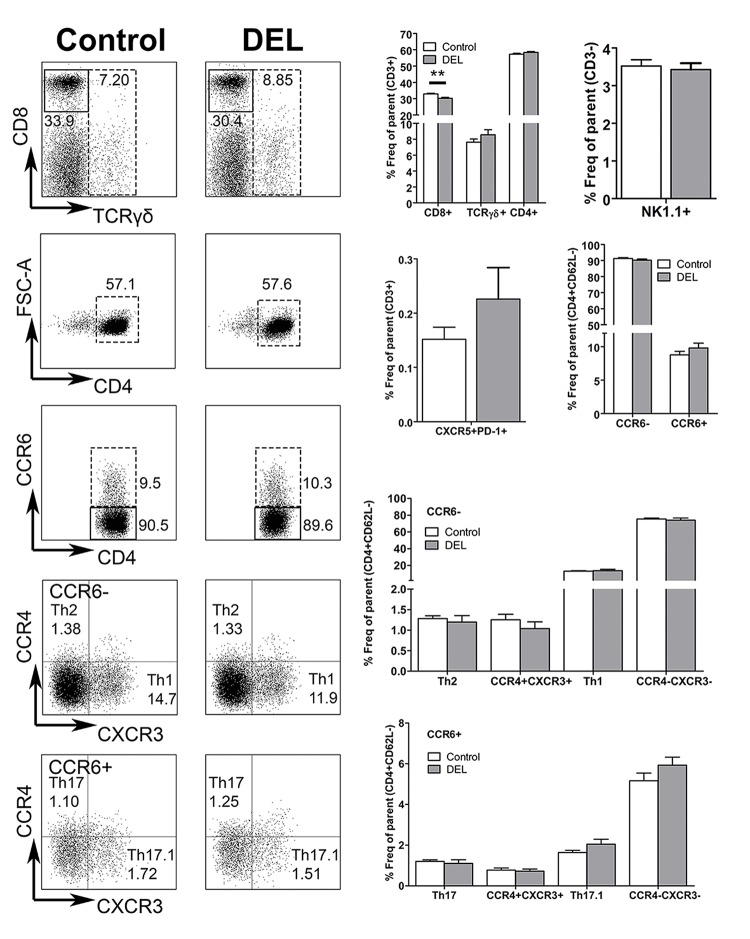
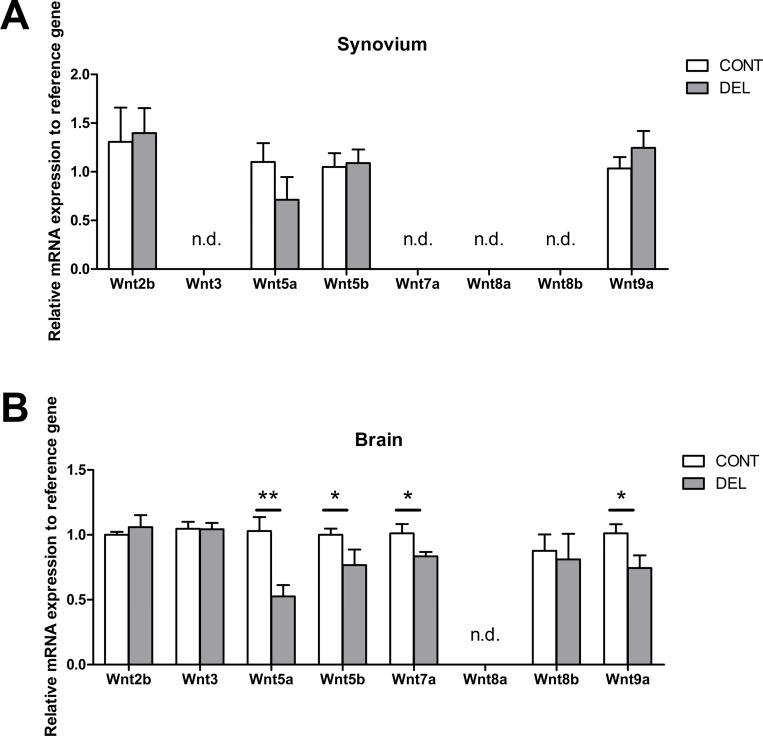
Dendritic cells (DCs) are professional antigen presenting cells that have the dual ability to stimulate immunity and maintain tolerance. However, the signalling pathways mediating tolerogenic DC function in vivo remain largely unknown. The β-catenin pathway has been suggested to promote a regulatory DC phenotype. The aim of this study was to unravel the role of β-catenin signalling to control DC function in the autoimmune collagen-induced arthritis model (CIA). Deletion of β-catenin specifically in DCs was achieved by crossing conditional knockout mice with a CD11c-Cre transgenic mouse line. Bone marrow-derived DCs (BMDCs) were generated and used to study the maturation profile of these cells in response to a TLR2 or TLR4 ligand stimulation. CIA was induced by intra-dermal immunization with 100 μg chicken type II collagen in complete Freund's adjuvant on days 0 and 21. CIA incidence and severity was monitored macroscopically and by histology. The T cell profile as well as their cytokine production were analysed by flow cytometry. Lack of β-catenin specifically in DCs did not affect the spontaneous, TLR2- or TLR4-induced maturation and activation of BMDCs or their cytokine production. Moreover, no effect on the incidence and severity of CIA was observed in mice lacking β-catenin in CD11c+ cells. A decreased frequency of splenic CD3+CD8+ T cells and of regulatory T cells (Tregs) (CD4+CD25highFoxP3+), but no changes in the frequency of splenic Th17 (CCR6+CXCR3-CCR4+), Th2 (CCR6-CXCR3-CCR4+) and Th1 (CCR6-CXCR3+CCR4-) cells were observed in these mice under CIA condition. Furthermore, the expression of IL-17A, IL-17F, IL-22, IL-4 or IFNγ was also not affected. Our data indicate that ablation of β-catenin expression in DCs did not alter the course and severity of CIA. We conclude that although deletion of β-catenin resulted in a lower frequency of Tregs, this decrease was not sufficient to aggravate the onset and severity of CIA.
Conflict of interest statement
Figures


References
-
- Takayanagi H. Osteoimmunology: shared mechanisms and crosstalk between the immune and bone systems. Nat Rev Immunol. 2007;7(4):292–304. - PubMed
-
- Steinman RM, Hawiger D, Liu K, Bonifaz L, Bonnyay D, Mahnke K, et al. Dendritic cell function in vivo during the steady state: a role in peripheral tolerance. Ann N Y Acad Sci. 2003;987:15–25. - PubMed
-
- Steinman RM, Hawiger D, Nussenzweig MC. Tolerogenic dendritic cells. Annu Rev Immunol. 2003;21:685–711. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
