

Contributions of Ca2+-Independent Thin Filament Activation to Cardiac Muscle Function
- PMID: 26588569
- PMCID: PMC4656859
- DOI: 10.1016/j.bpj.2015.09.028
Contributions of Ca2+-Independent Thin Filament Activation to Cardiac Muscle Function
Abstract
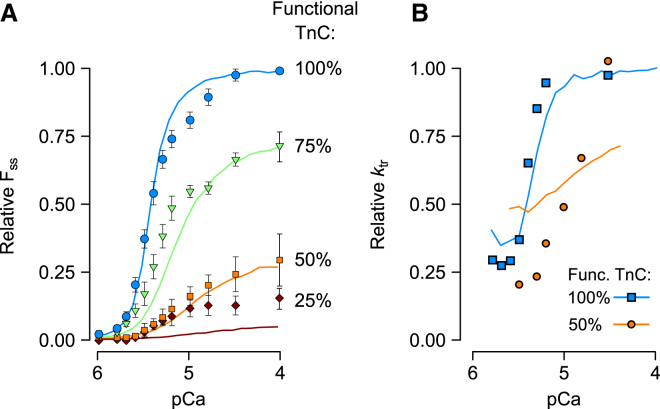
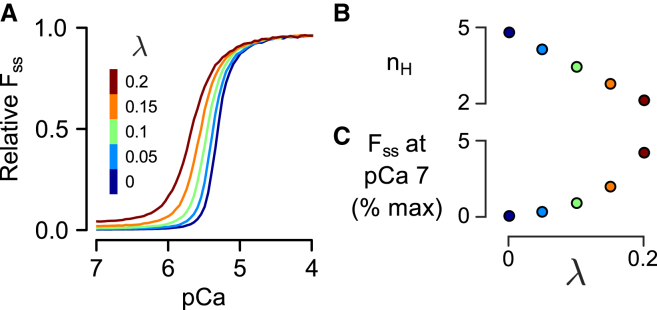
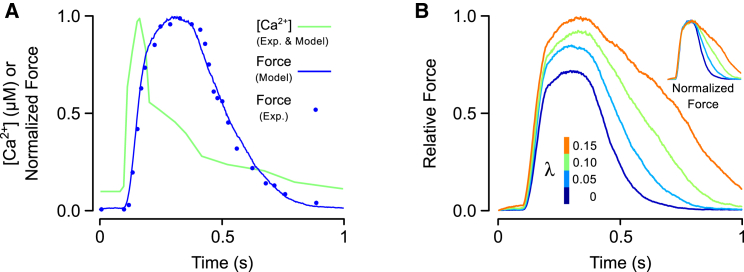
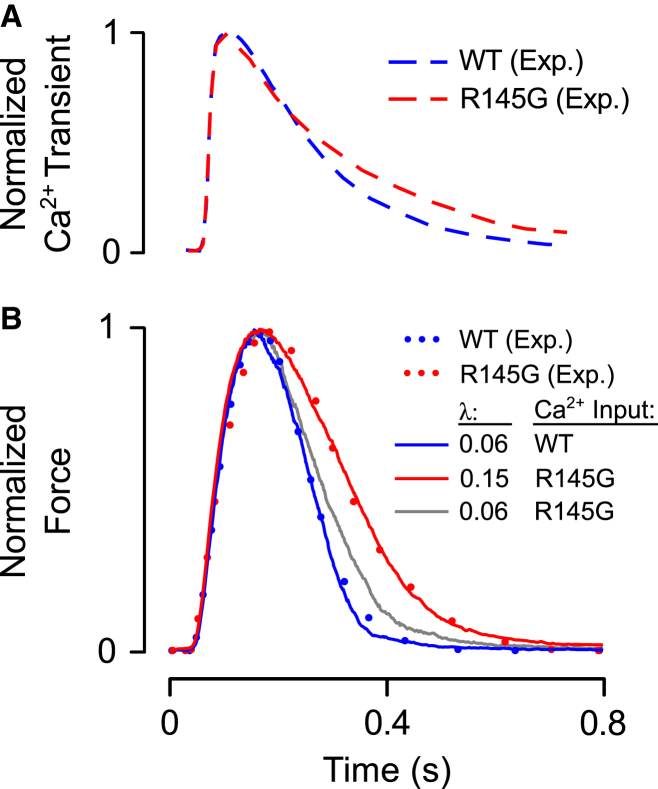
Although Ca2+ is the principal regulator of contraction in striated muscle, in vitro evidence suggests that some actin-myosin interaction is still possible even in its absence. Whether this Ca2+-independent activation (CIA) occurs under physiological conditions remains unclear, as does its potential impact on the function of intact cardiac muscle. The purpose of this study was to investigate CIA using computational analysis. We added a structurally motivated representation of this phenomenon to an existing myofilament model, which allowed predictions of CIA-dependent muscle behavior. We found that a certain amount of CIA was essential for the model to reproduce reported effects of nonfunctional troponin C on myofilament force generation. Consequently, those data enabled estimation of ΔGCIA, the energy barrier for activating a thin filament regulatory unit in the absence of Ca2+. Using this estimate of ΔGCIA as a point of reference (∼7 kJ mol(-1)), we examined its impact on various aspects of muscle function through additional simulations. CIA decreased the Hill coefficient of steady-state force while increasing myofilament Ca2+ sensitivity. At the same time, CIA had minimal effect on the rate of force redevelopment after slack/restretch. Simulations of twitch tension show that the presence of CIA increases peak tension while profoundly delaying relaxation. We tested the model's ability to represent perturbations to the Ca2+ regulatory mechanism by analyzing twitch records measured in transgenic mice expressing a cardiac troponin I mutation (R145G). The effects of the mutation on twitch dynamics were fully reproduced by a single parameter change, namely lowering ΔGCIA by 2.3 kJ mol(-1) relative to its wild-type value. Our analyses suggest that CIA is present in cardiac muscle under normal conditions and that its modulation by gene mutations or other factors can alter both systolic and diastolic function.
Copyright © 2015 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
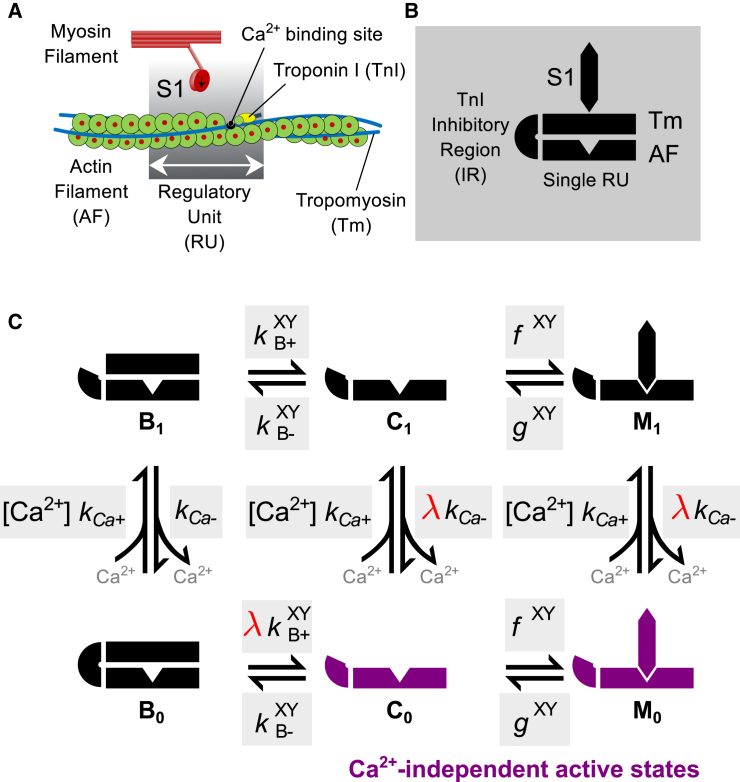
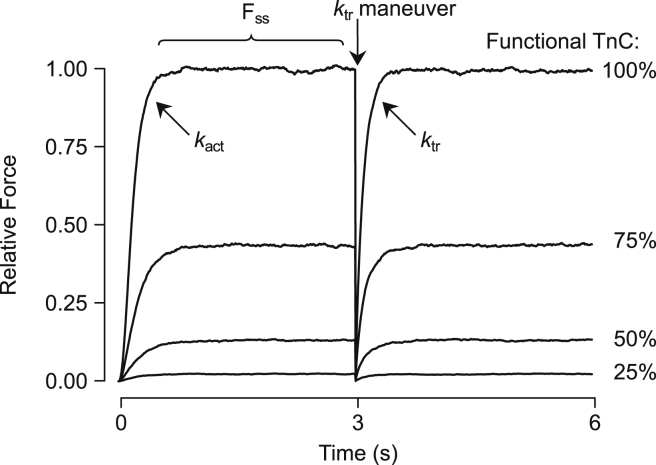
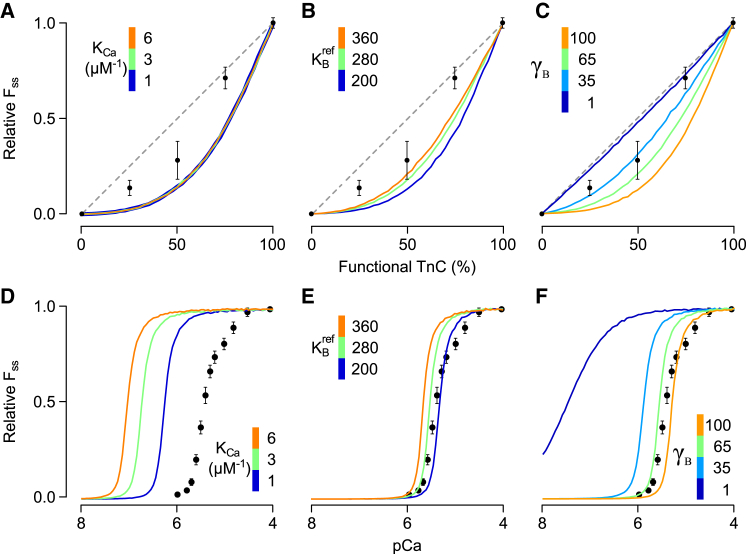
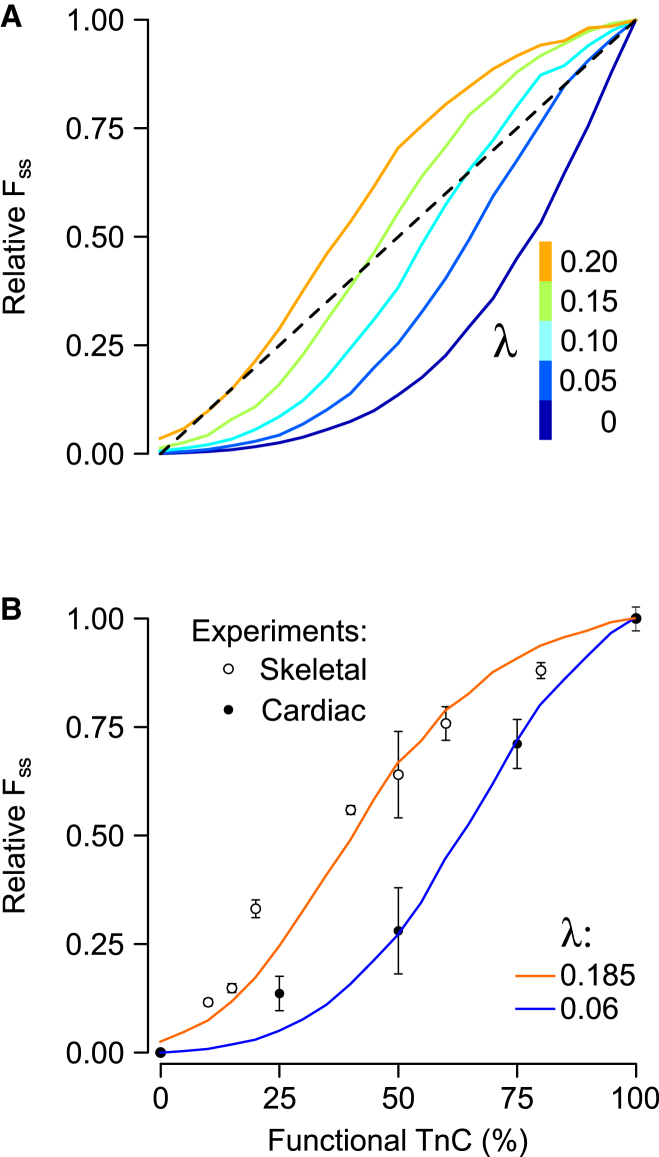
References
-
- Gordon A.M., Homsher E., Regnier M. Regulation of contraction in striated muscle. Physiol. Rev. 2000;80:853–924. - PubMed
-
- Fuchs F. Thermal inactivation of the calcium regulatory mechanism of human skeletal muscle actomyosin: a possible contributing factor in the rigidity of malignant hyperthermia. Anesthesiology. 1975;42:584–589. - PubMed
-
- Schaertl S., Lehrer S.S., Geeves M.A. Separation and characterization of the two functional regions of troponin involved in muscle thin filament regulation. Biochemistry. 1995;34:15890–15894. - PubMed
-
- Maytum R., Westerdorf B., Geeves M.A. Differential regulation of the actomyosin interaction by skeletal and cardiac troponin isoforms. J. Biol. Chem. 2003;278:6696–6701. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
