

Identification of Common Features in Prototype Broadly Neutralizing Antibodies to HIV Envelope V2 Apex to Facilitate Vaccine Design
- PMID: 26588781
- PMCID: PMC4654981
- DOI: 10.1016/j.immuni.2015.10.014
Identification of Common Features in Prototype Broadly Neutralizing Antibodies to HIV Envelope V2 Apex to Facilitate Vaccine Design
Abstract
Broadly neutralizing antibodies (bnAbs) directed to the V2 apex of the HIV envelope (Env) trimer isolated from individual HIV-infected donors potently neutralize diverse HIV strains, but strategies for designing immunogens to elicit bnAbs have not been identified. Here, we compared four prototypes (PG9, CH01, PGT145, and CAP256.VRC26.09) of V2 apex bnAbs and showed that all recognized a core epitope of basic V2 residues and the glycan-N160. Two prototype bnAbs were derived from VH-germlines that were 99% identical and used a common germline D-gene encoded YYD-motif to interact with the V2-epitope. We identified isolates that were neutralized by inferred germline (iGL) versions of three of the prototype bnAbs. Soluble Env derived from one of these isolates was shown to form a well-ordered Env trimer that could serve as an immunogen to initiate a V2-apex bnAb response. These studies illustrate a strategy to transition from panels of bnAbs to vaccine candidates.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
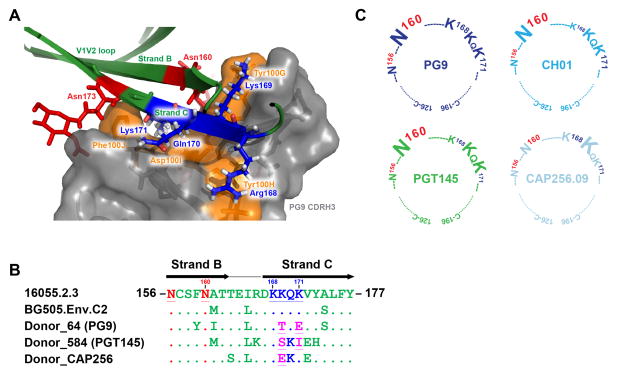



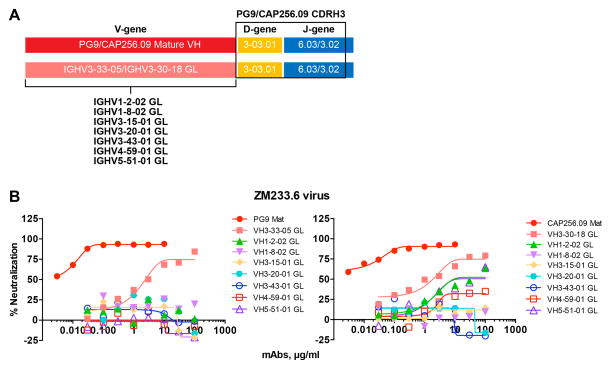

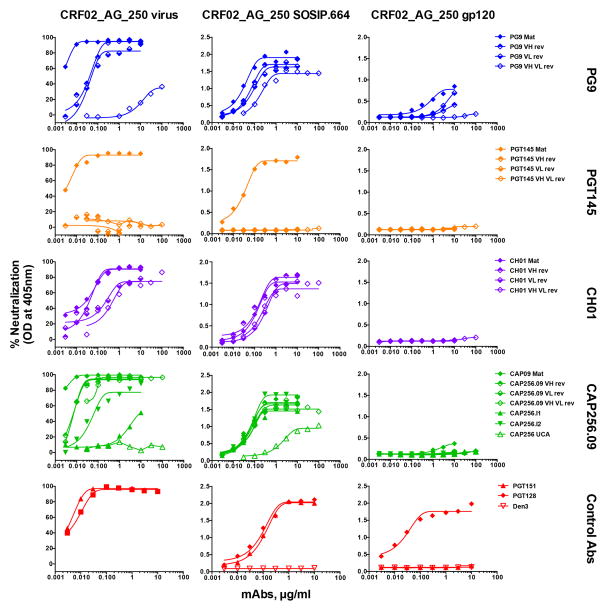
Comment in
-
Common Characteristics of HIV-Neutralizing Antibodies with a Fondness for Sugars.Immunity. 2015 Nov 17;43(5):837-40. doi: 10.1016/j.immuni.2015.10.026. Immunity. 2015. PMID: 26588775
References
-
- Bonsignori M, Hwang KK, Chen X, Tsao CY, Morris L, Gray E, Marshall DJ, Crump JA, Kapiga SH, Sam NE, et al. Analysis of a clonal lineage of HIV-1 envelope V2/V3 conformational epitope-specific broadly neutralizing antibodies and their inferred unmutated common ancestors. Journal of virology. 2011;85:9998–10009. - PMC - PubMed
-
- Braibant M, Gong EY, Plantier JC, Moreau T, Alessandri E, Simon F, Barin F. Cross-group neutralization of HIV-1 and evidence for conservation of the PG9/PG16 epitopes within divergent groups. Aids. 2013;27:1239–1244. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
