

Morphometrics, 3D Imaging, and Craniofacial Development
- PMID: 26589938
- PMCID: PMC5299999
- DOI: 10.1016/bs.ctdb.2015.09.003
Morphometrics, 3D Imaging, and Craniofacial Development
Abstract
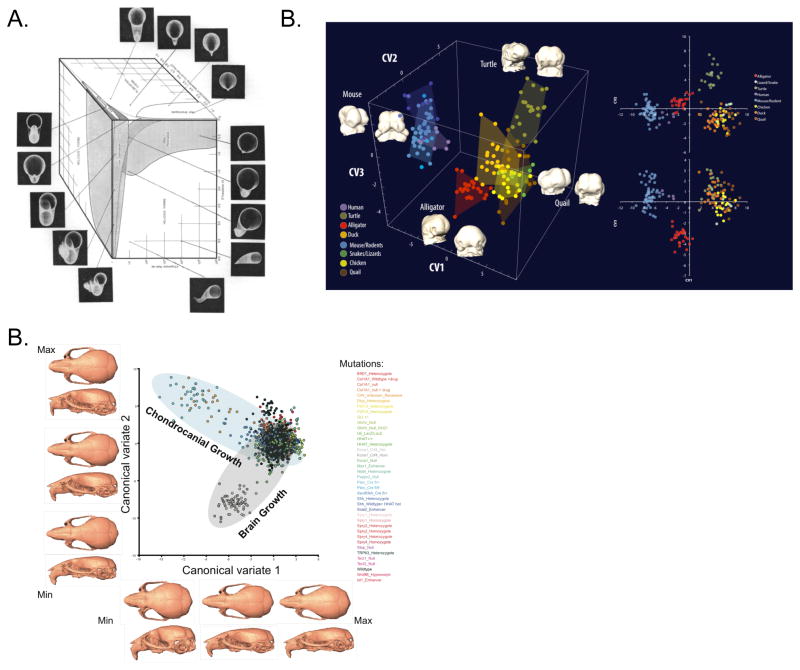
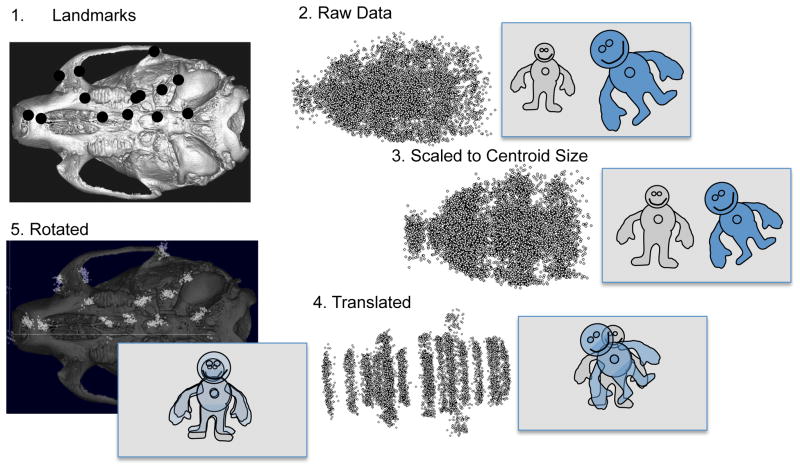
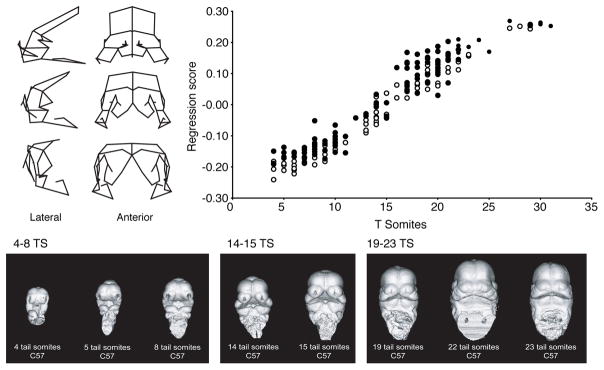
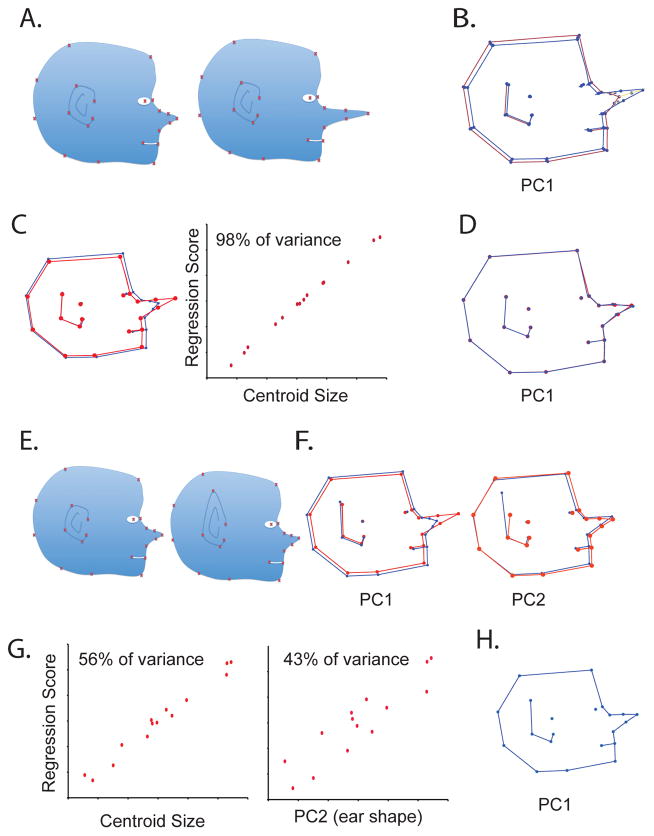
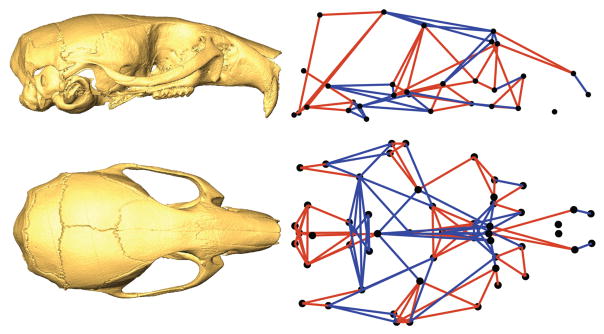
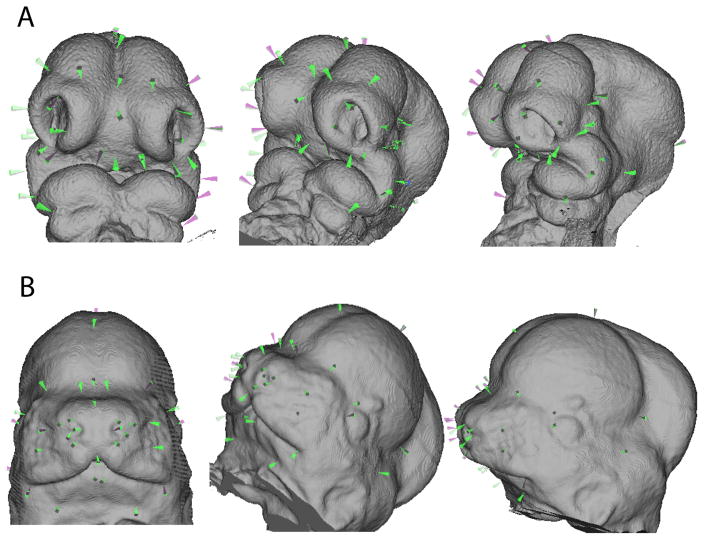
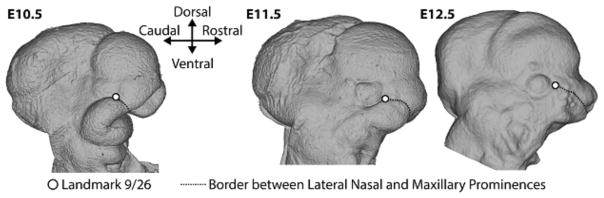
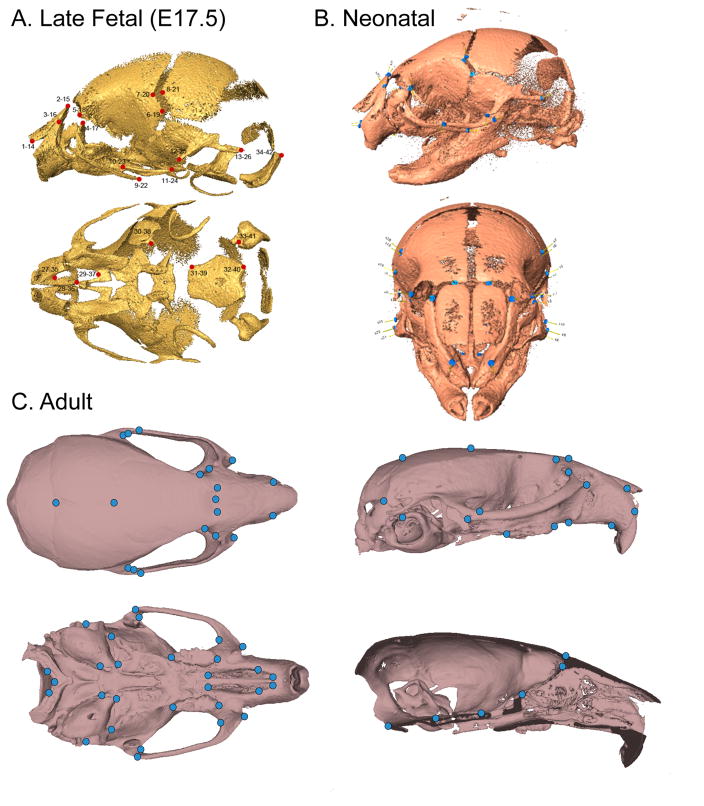
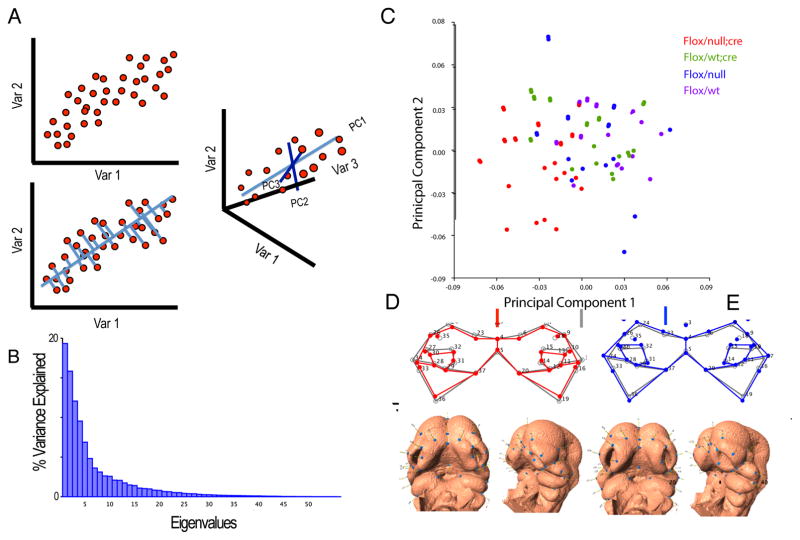
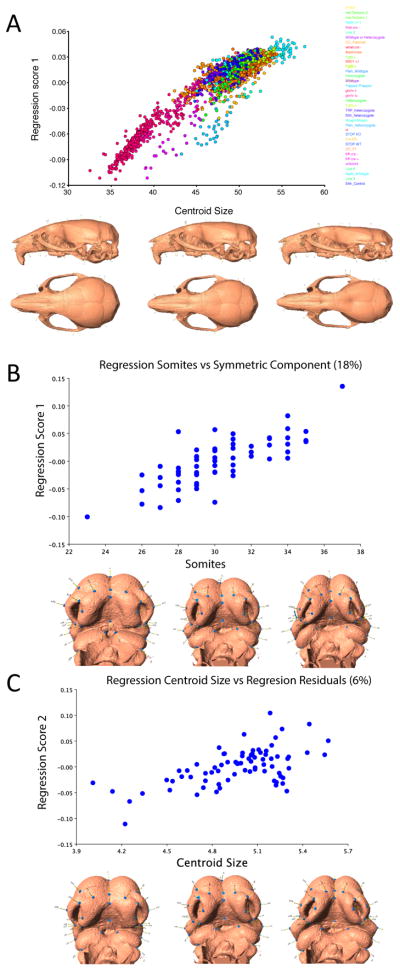
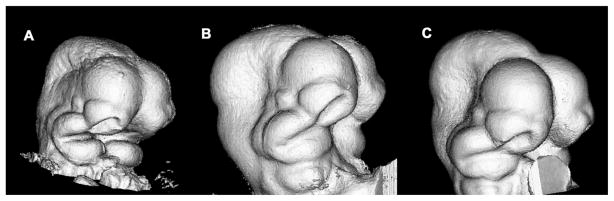
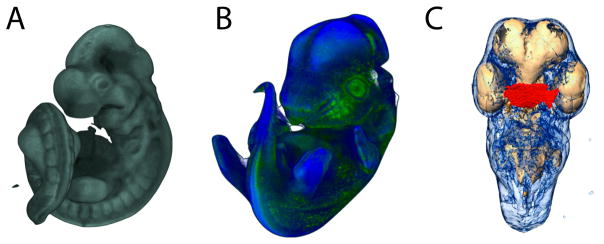
Recent studies have shown how volumetric imaging and morphometrics can add significantly to our understanding of morphogenesis, the developmental basis for variation, and the etiology of structural birth defects. On the other hand, the complex questions and diverse imaging data in developmental biology present morphometrics with more complex challenges than applications in virtually any other field. Meeting these challenges is necessary in order to understand the mechanistic basis for variation in complex morphologies. This chapter reviews the methods and theory that enable the application of modern landmark-based morphometrics to developmental biology and craniofacial development, in particular. We discuss the theoretical foundations of morphometrics as applied to development and review the basic approaches to the quantification of morphology. Focusing on geometric morphometrics, we discuss the principal statistical methods for quantifying and comparing morphological variation and covariation structure within and among groups. Finally, we discuss the future directions for morphometrics in developmental biology that will be required for approaches that enable quantitative integration across the genotype-phenotype map.
Keywords: Craniofacial; Imaging; MicroCT; Morphogenesis; Morphometrics; Mouse; Optical Projection Tomography; Phenomics.
© 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Alberch P. Developmental constraints in evolutionary processes. In: Bonner JT, editor. Development in Evolution. Berlin and New York: Springer-Verlag; 1982. pp. 313–332.
-
- Bastir M, Rosas A. Hierarchical nature of morphological integration and modularity in the human posterior face. Am J Phys Anthropol. 2005;128(1):26–34. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
