

Warts phosphorylates mud to promote pins-mediated mitotic spindle orientation in Drosophila, independent of Yorkie
- PMID: 26592339
- PMCID: PMC4660254
- DOI: 10.1016/j.cub.2015.09.025
Warts phosphorylates mud to promote pins-mediated mitotic spindle orientation in Drosophila, independent of Yorkie
Abstract
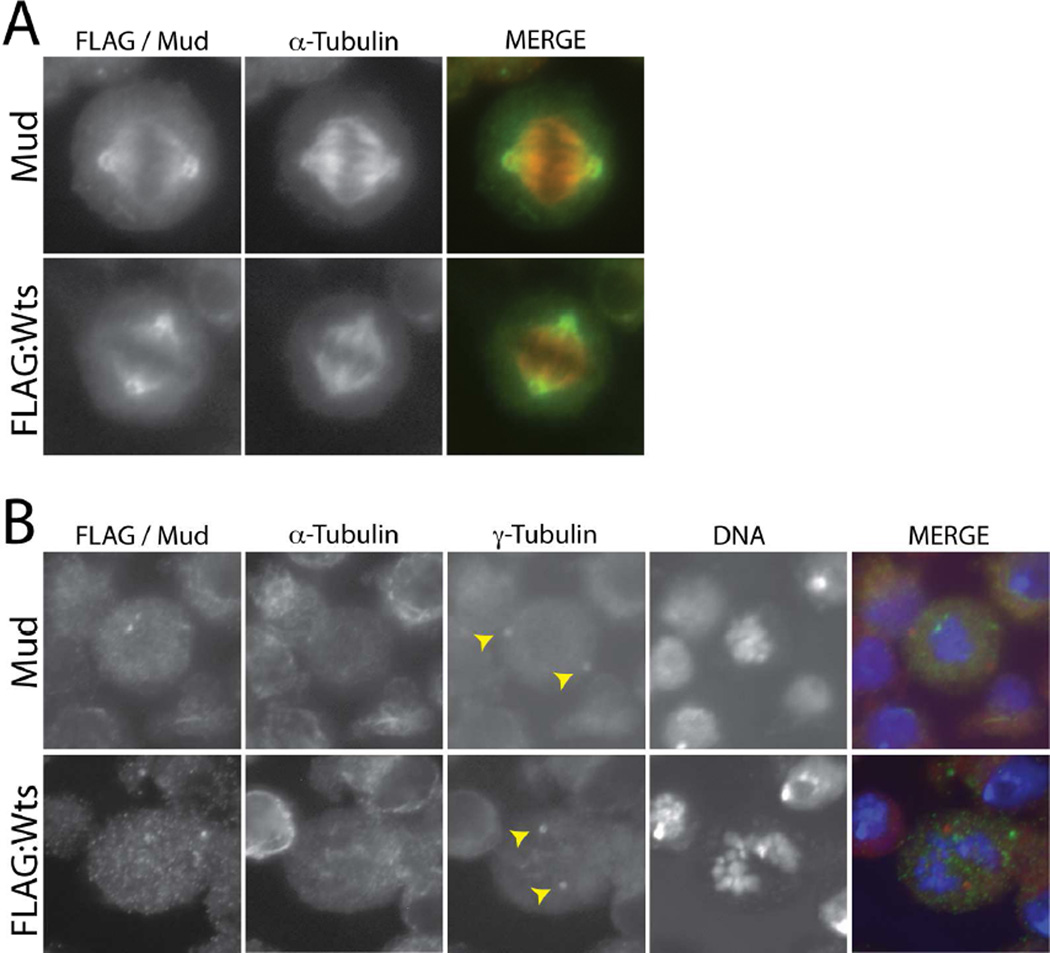
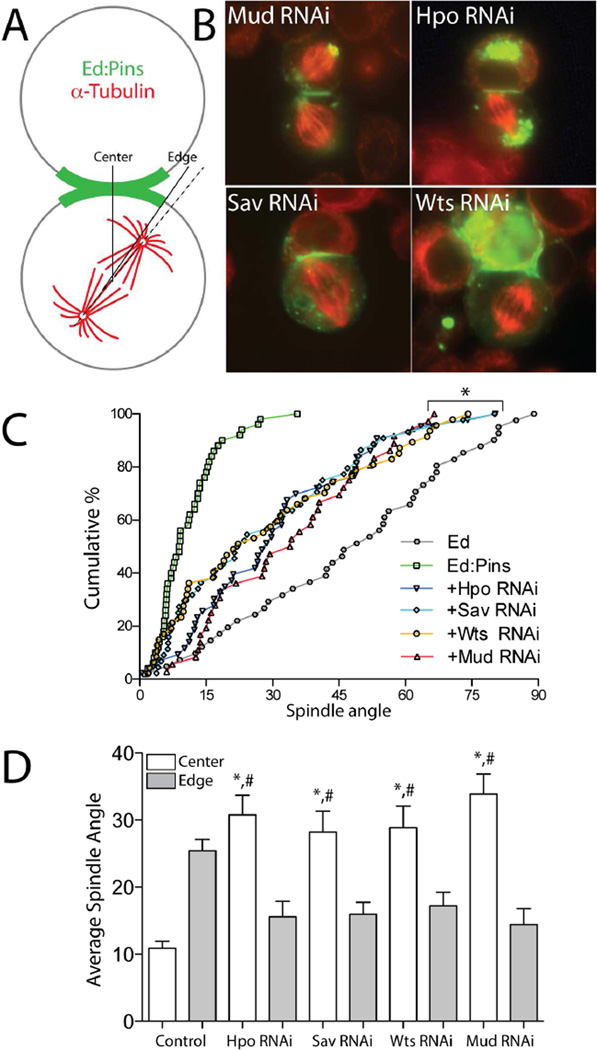
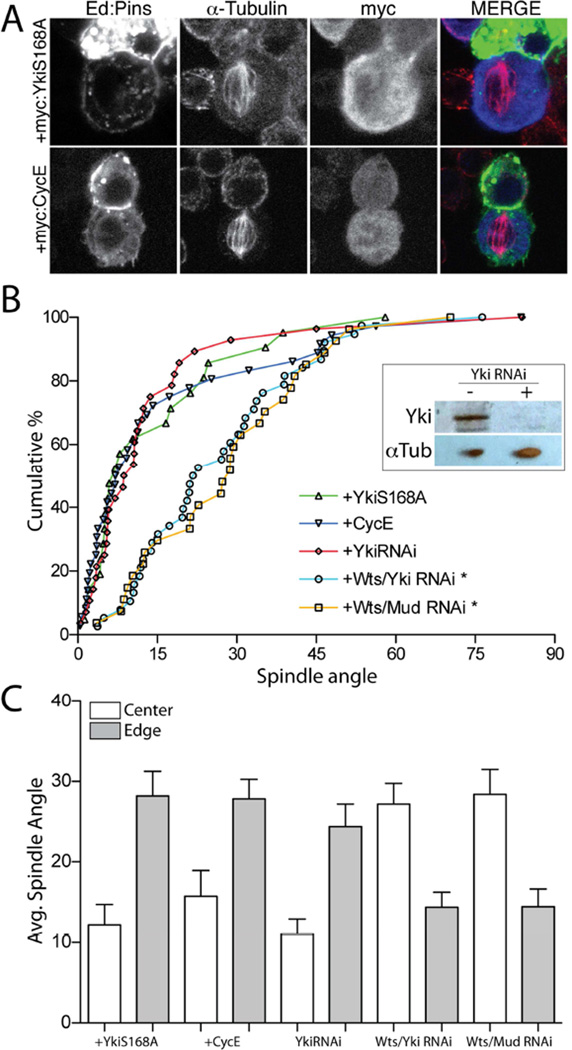
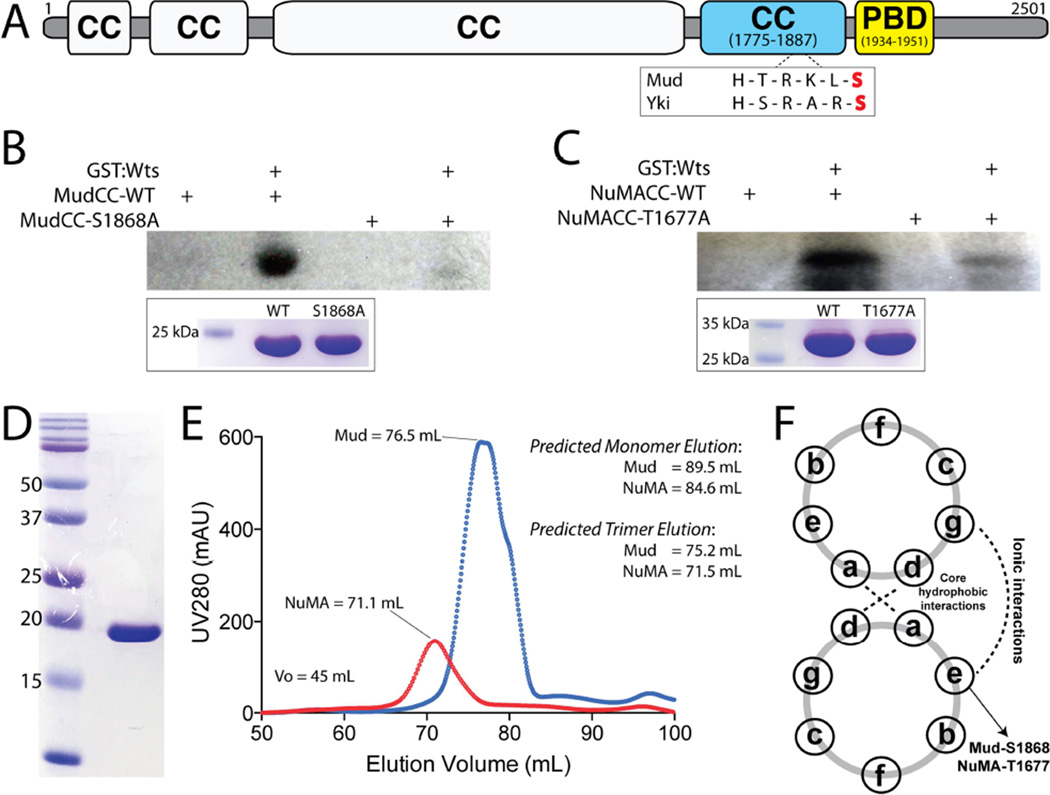
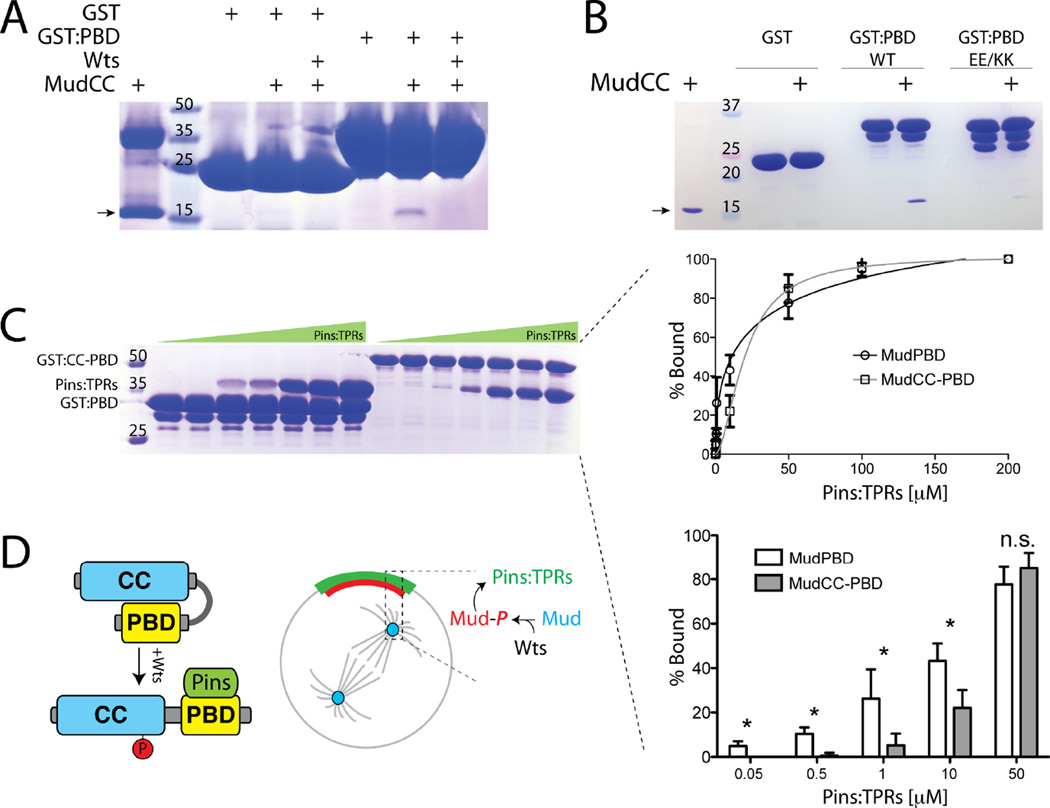
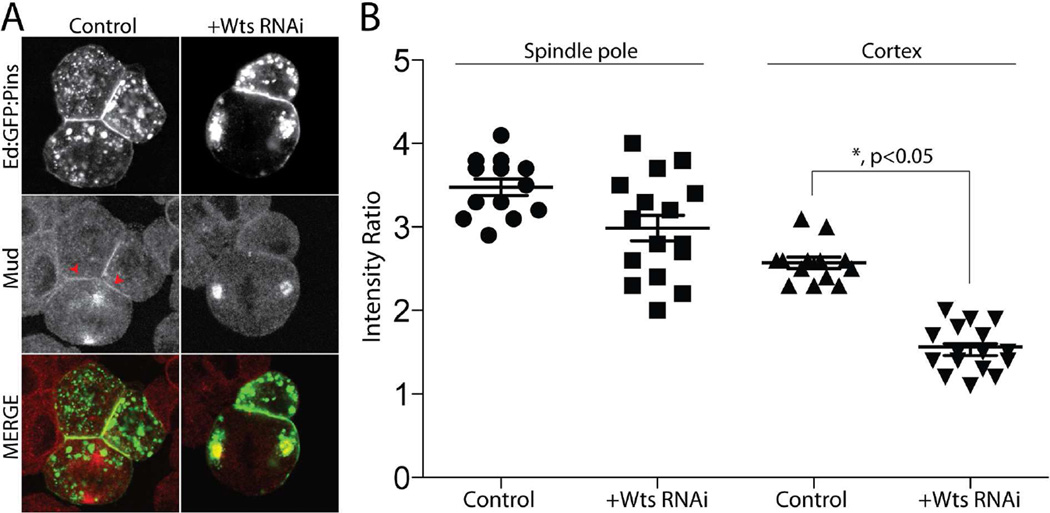
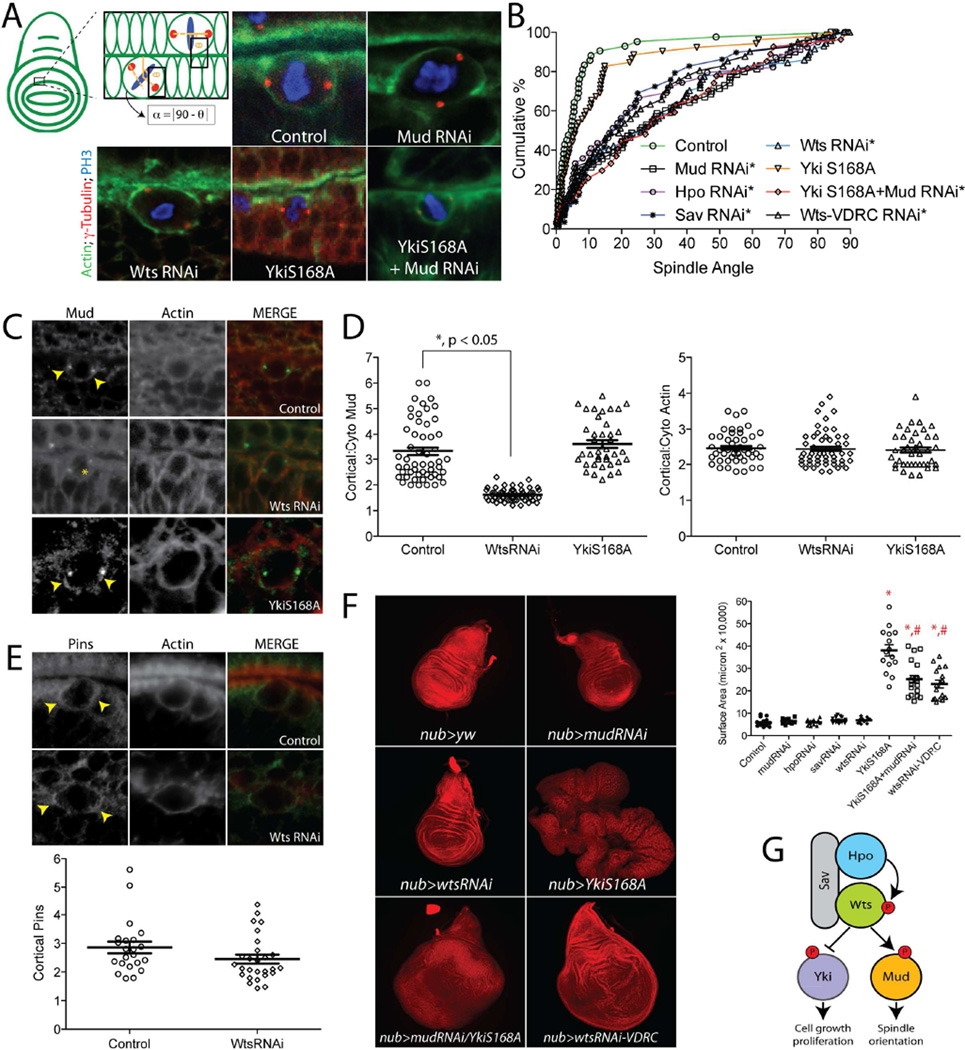
Multicellular animals have evolved conserved signaling pathways that translate cell polarity cues into mitotic spindle positioning to control the orientation of cell division within complex tissue structures. These oriented cell divisions are essential for the development of cell diversity and the maintenance of tissue homeostasis. Despite intense efforts, the molecular mechanisms that control spindle orientation remain incompletely defined. Here, we describe a role for the Hippo (Hpo) kinase complex in promoting Partner of Inscuteable (Pins)-mediated spindle orientation. Knockdown of Hpo, Salvador (Sav), or Warts (Wts) each result in a partial loss of spindle orientation, a phenotype previously described following loss of the Pins-binding protein Mushroom body defect (Mud). Similar to orthologs spanning yeast to mammals, Wts kinase localizes to mitotic spindle poles, a prominent site of Mud localization. Wts directly phosphorylates Mud in vitro within its C-terminal coiled-coil domain. This Mud coiled-coil domain directly binds the adjacent Pins-binding domain to dampen the Pins/Mud interaction, and Wts-mediated phosphorylation uncouples this intramolecular Mud interaction. Loss of Wts prevents cortical Pins/Mud association without affecting Mud accumulation at spindle poles, suggesting phosphorylation acts as a molecular switch to specifically activate cortical Mud function. Finally, loss of Wts in Drosophila imaginal disc epithelial cells results in diminished cortical Mud and defective planar spindle orientation. Our results provide new insights into the molecular basis for dynamic regulation of the cortical Pins/Mud spindle positioning complex and highlight a novel link with an essential, evolutionarily conserved cell proliferation pathway.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- Cabernard C, Doe CQ. Apical/basal spindle orientation is required for neuroblast homeostasis and neuronal differentiation in Drosophila. Developmental cell. 2009;17:134–141. - PubMed
-
- Gonzalez C. Spindle orientation, asymmetric division and tumour suppression in Drosophila stem cells. Nature reviews. Genetics. 2007;8:462–472. - PubMed
-
- Bellaiche Y, Radovic A, Woods DF, Hough CD, Parmentier ML, O'Kane CJ, Bryant PJ, Schweisguth F. The Partner of Inscuteable/Discs-large complex is required to establish planar polarity during asymmetric cell division in Drosophila. Cell. 2001;106:355–366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
