

Structure of human carbamoyl phosphate synthetase: deciphering the on/off switch of human ureagenesis
- PMID: 26592762
- PMCID: PMC4655335
- DOI: 10.1038/srep16950
Structure of human carbamoyl phosphate synthetase: deciphering the on/off switch of human ureagenesis
Abstract
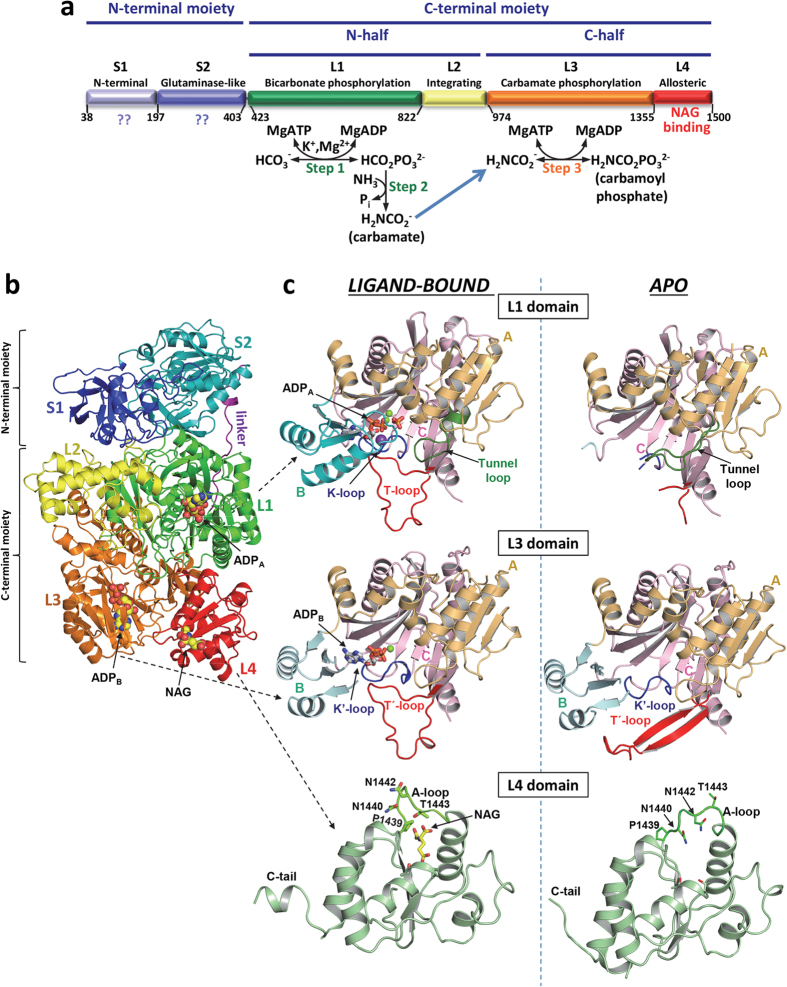
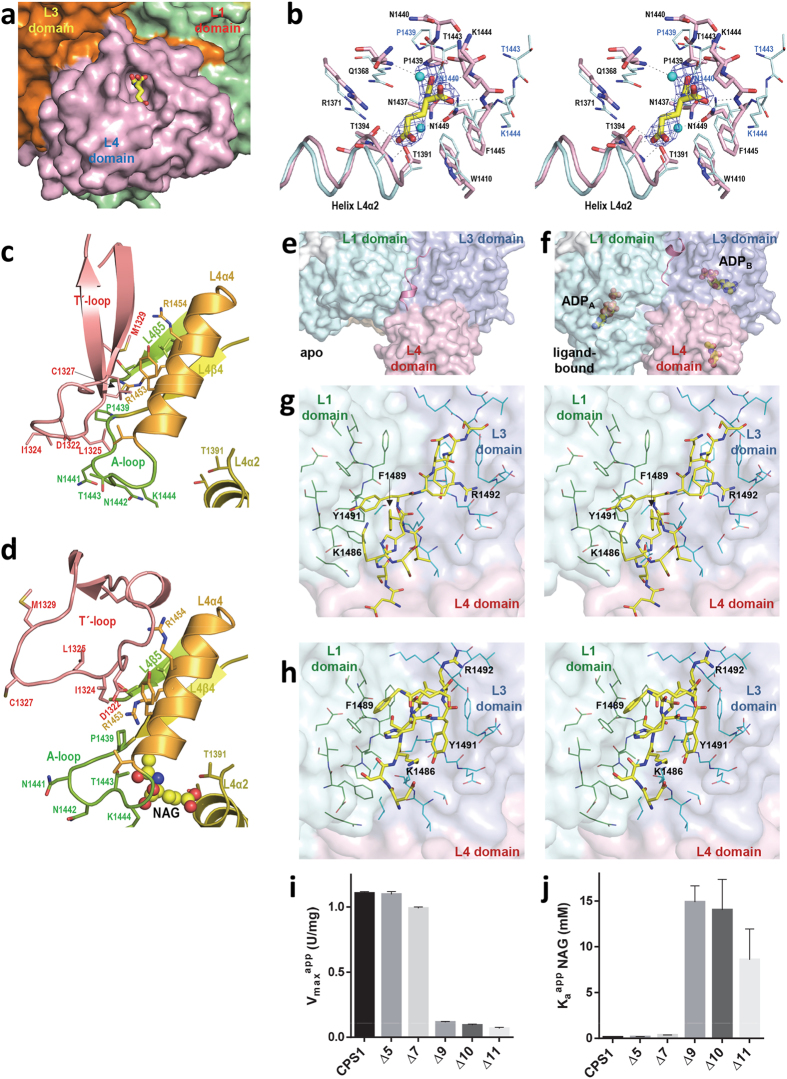
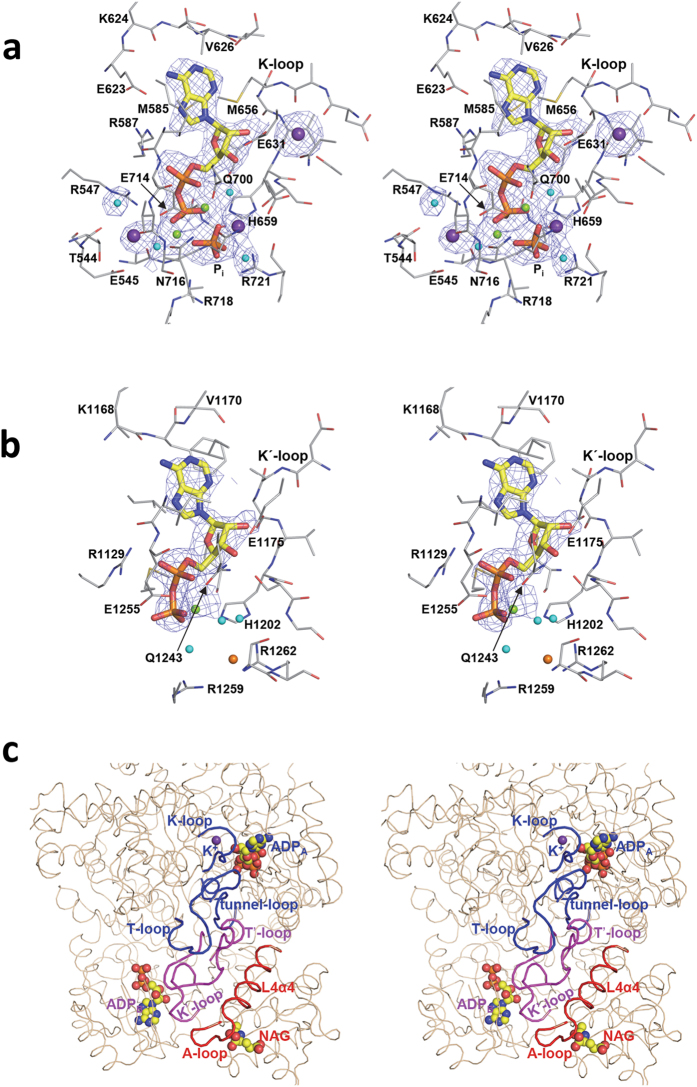
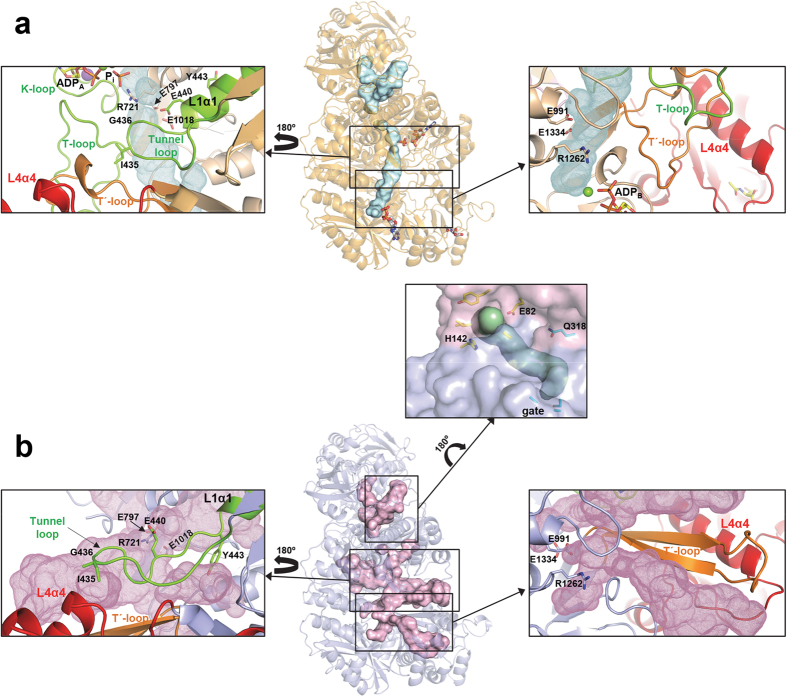
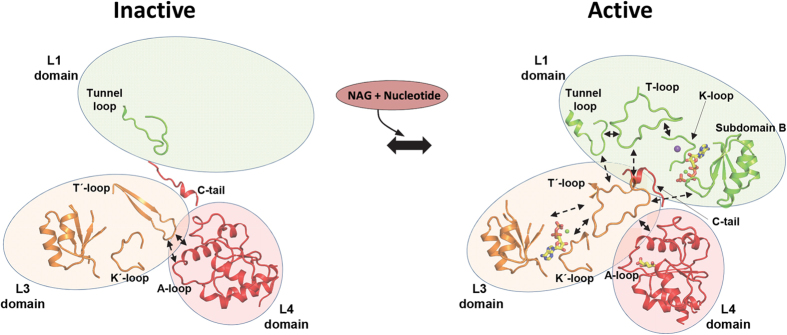
Human carbamoyl phosphate synthetase (CPS1), a 1500-residue multidomain enzyme, catalyzes the first step of ammonia detoxification to urea requiring N-acetyl-L-glutamate (NAG) as essential activator to prevent ammonia/amino acids depletion. Here we present the crystal structures of CPS1 in the absence and in the presence of NAG, clarifying the on/off-switching of the urea cycle by NAG. By binding at the C-terminal domain of CPS1, NAG triggers long-range conformational changes affecting the two distant phosphorylation domains. These changes, concerted with the binding of nucleotides, result in a dramatic remodeling that stabilizes the catalytically competent conformation and the building of the ~35 Å-long tunnel that allows migration of the carbamate intermediate from its site of formation to the second phosphorylation site, where carbamoyl phosphate is produced. These structures allow rationalizing the effects of mutations found in patients with CPS1 deficiency (presenting hyperammonemia, mental retardation and even death), as exemplified here for some mutations.
Figures
References
-
- Cohen P. P., Hall L. M. & Metzenberg R. L. Isolation and characterization of a naturally occurring stimulator of citrulline biosynthesis. Nature 178, 1468–1469 (1956). - PubMed
-
- Rubio V., Ramponi G. & Grisolia S. Carbamoyl phosphate synthetase I of human liver. Purification, some properties and immunological cross-reactivity with the rat liver enzyme. Biochim. Biophys. Acta, 659, 150–160 (1981). - PubMed
-
- Bachmann C., Colombo J. P. & Jaggi K. N-acetylglutamate synthetase (NAGS) deficiency: diagnosis, clinical observations and treatment. Adv. Exp. Med. Biol. 153, 39–45 (1982). - PubMed
-
- Brusilow S. W. & Horwich A. L. Urea cycle enzymes. In Scriver C. R., Beaudet A. L., Sly W. S. & Valle D., editors; Child, B., Kinzler, K. W. & Vogelstein, B., associated editors. The Metabolic and Molecular Bases of Inherited Disease. 8th edition. New York, McGraw-Hill. Vol 2, pp 1909–1963 (2001).
-
- Stewart P. M. & Walser M. Short term regulation of ureagenesis. J. Biol. Chem. 255, 5270–5280 (1980). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
