

Glutamine synthetase activity fuels nucleotide biosynthesis and supports growth of glutamine-restricted glioblastoma
- PMID: 26595383
- PMCID: PMC4663685
- DOI: 10.1038/ncb3272
Glutamine synthetase activity fuels nucleotide biosynthesis and supports growth of glutamine-restricted glioblastoma
Abstract
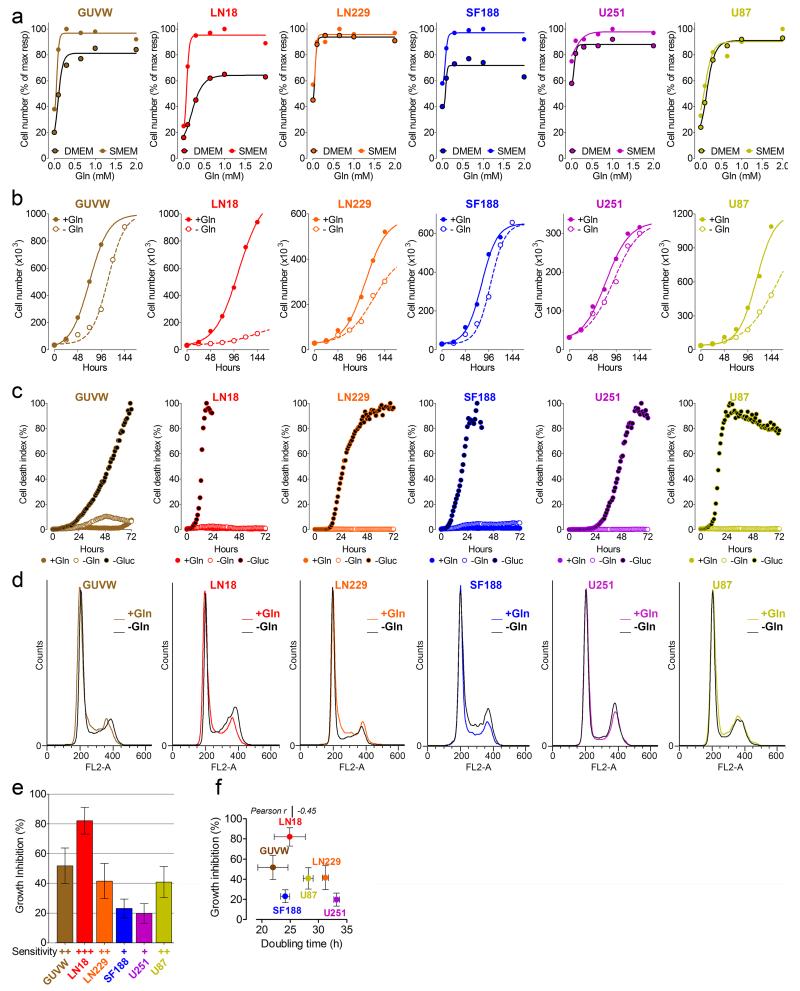
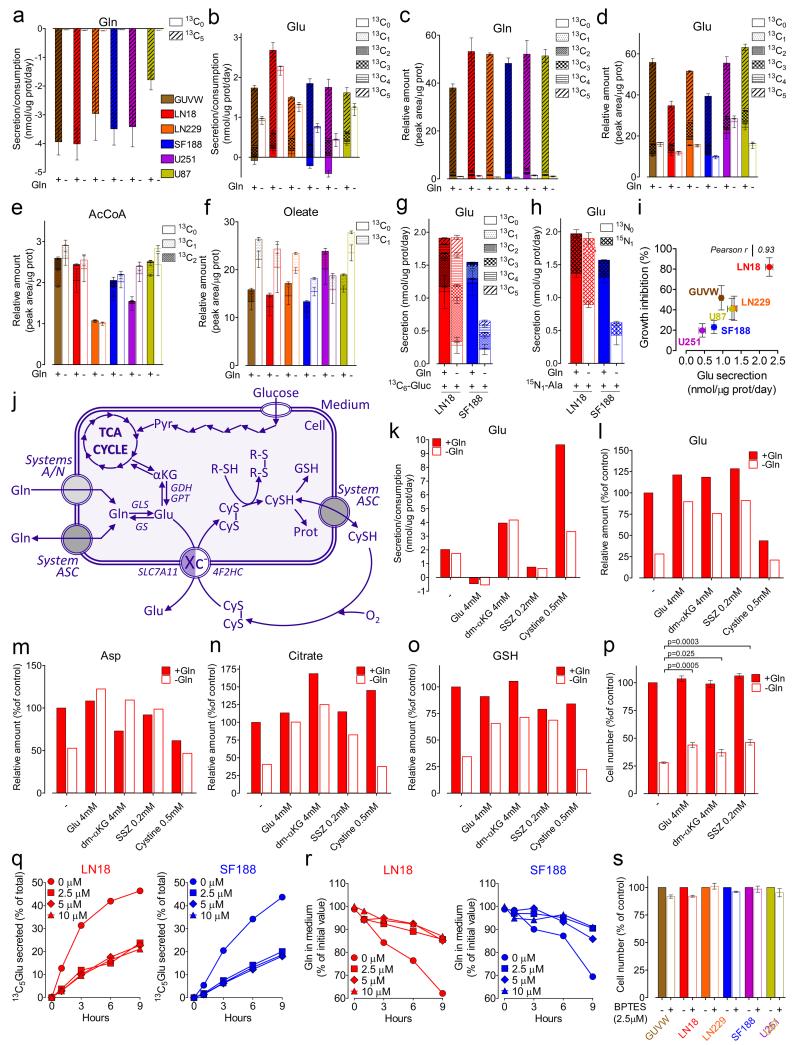
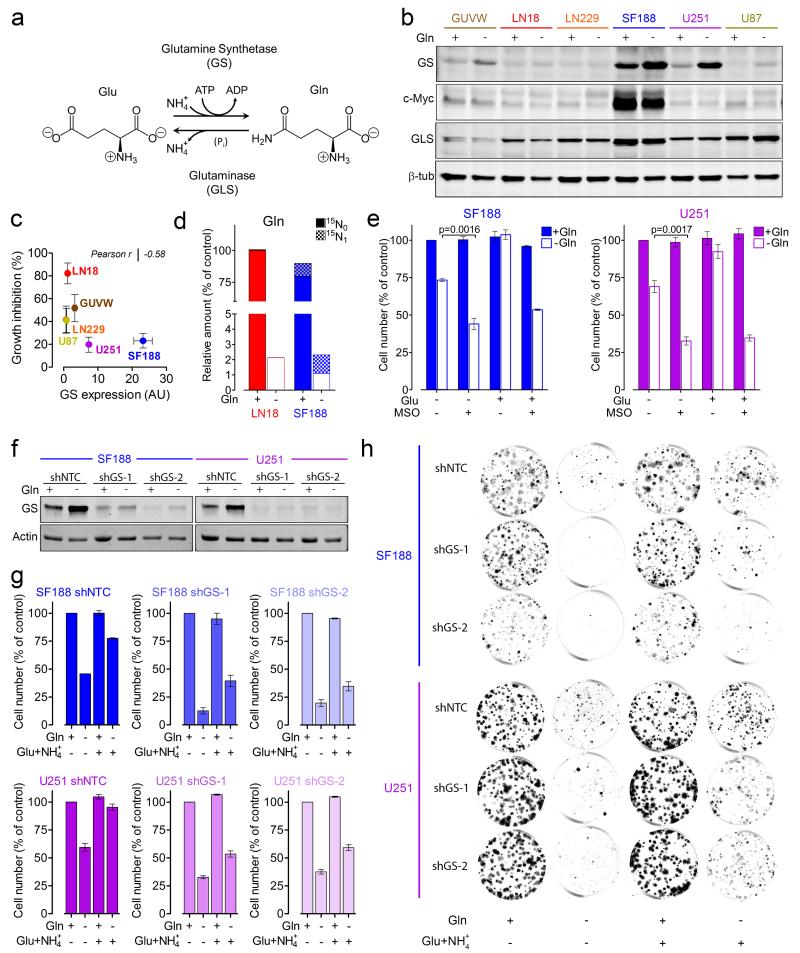
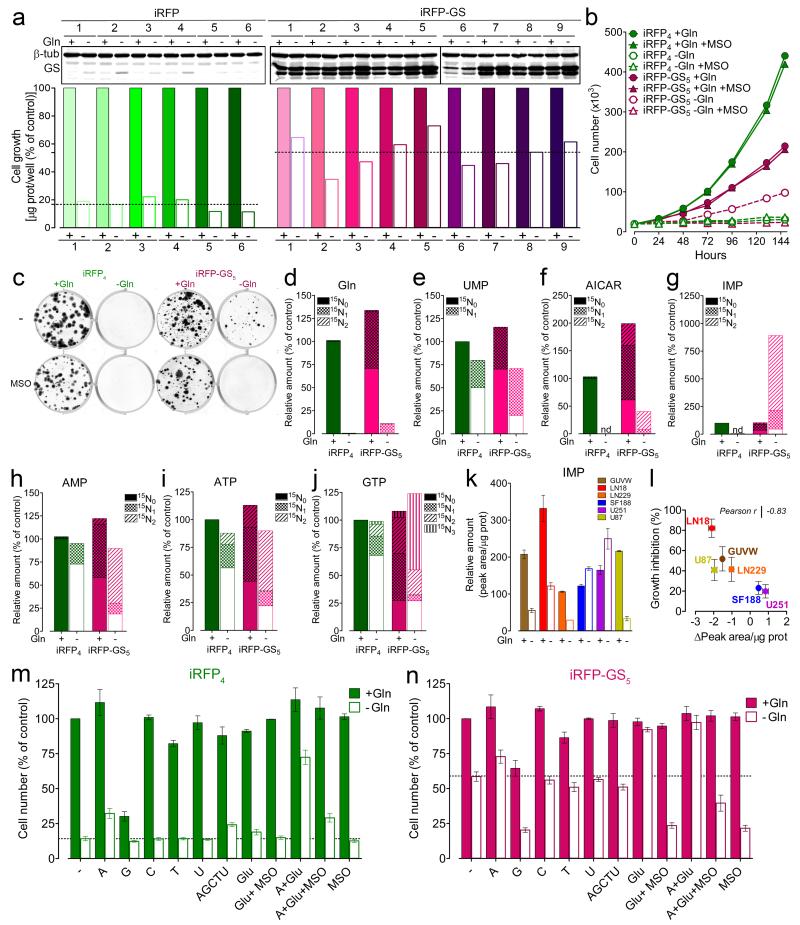
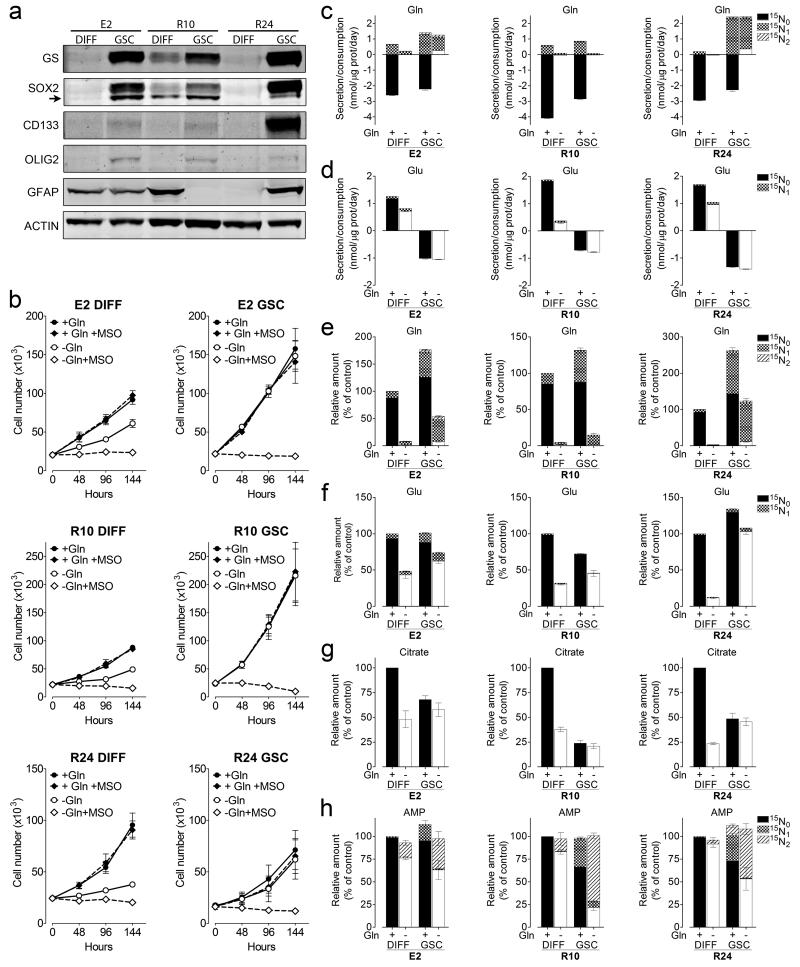
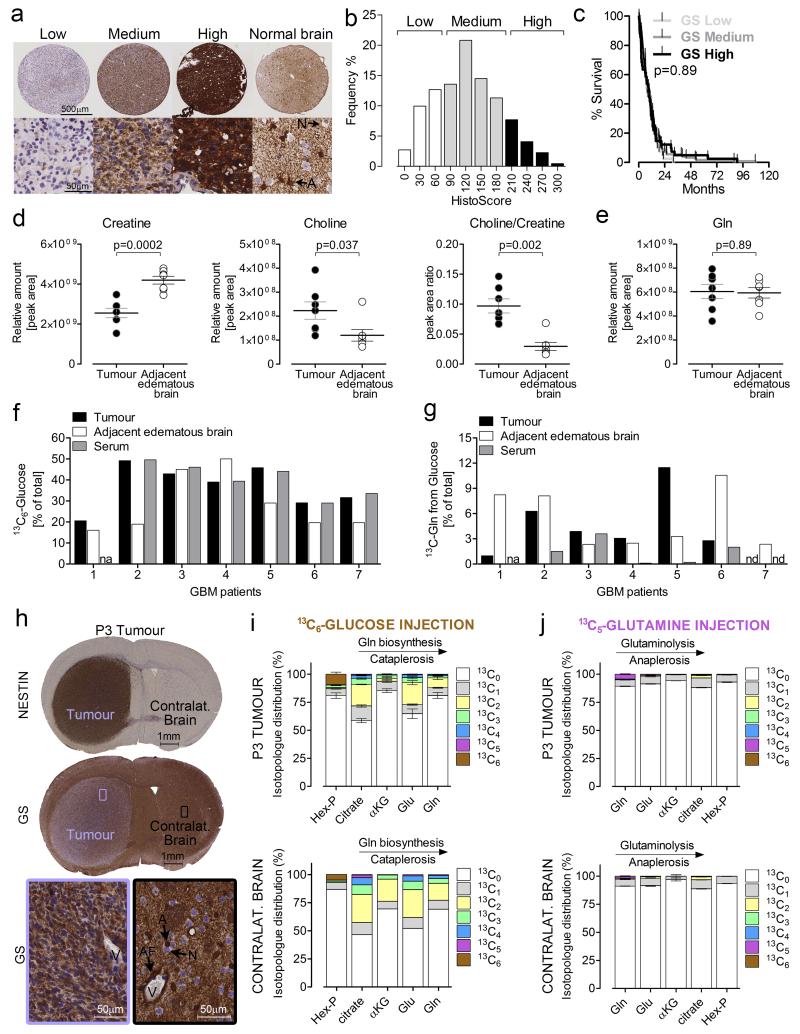
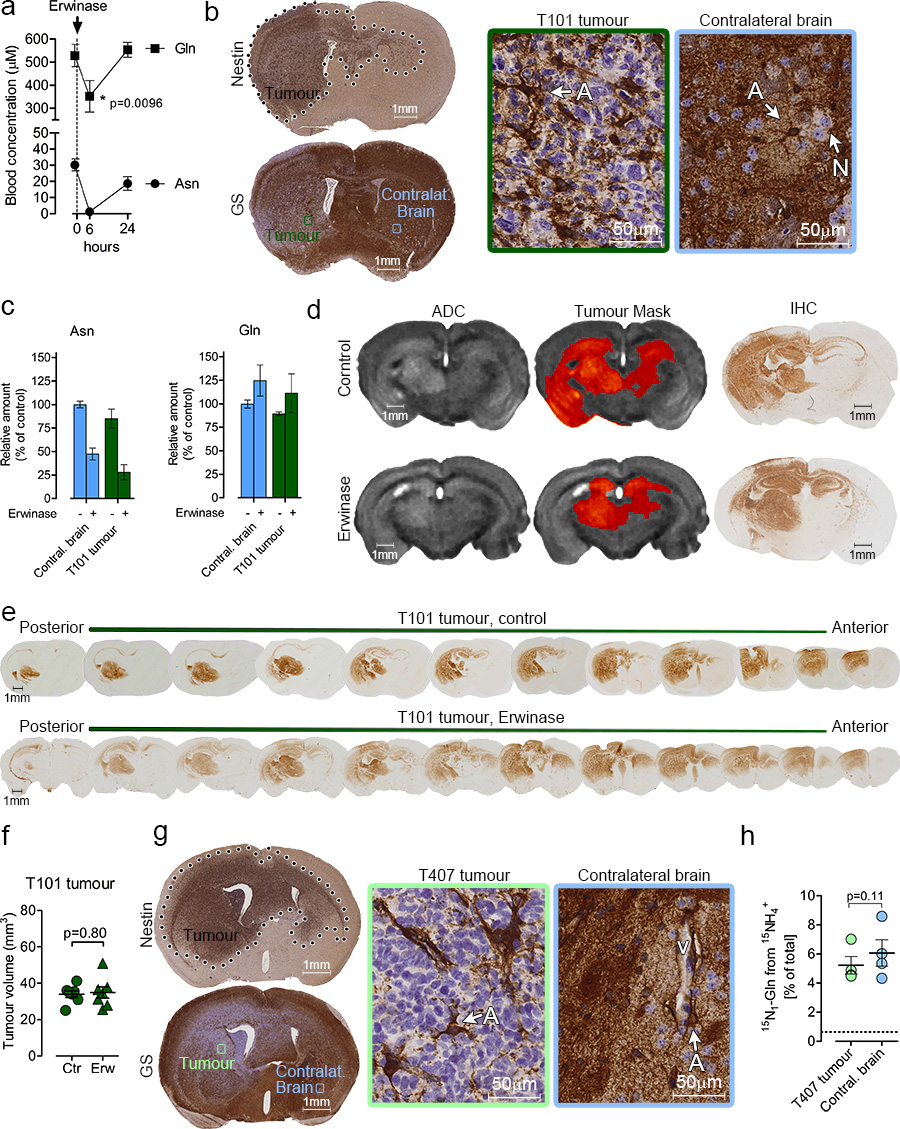
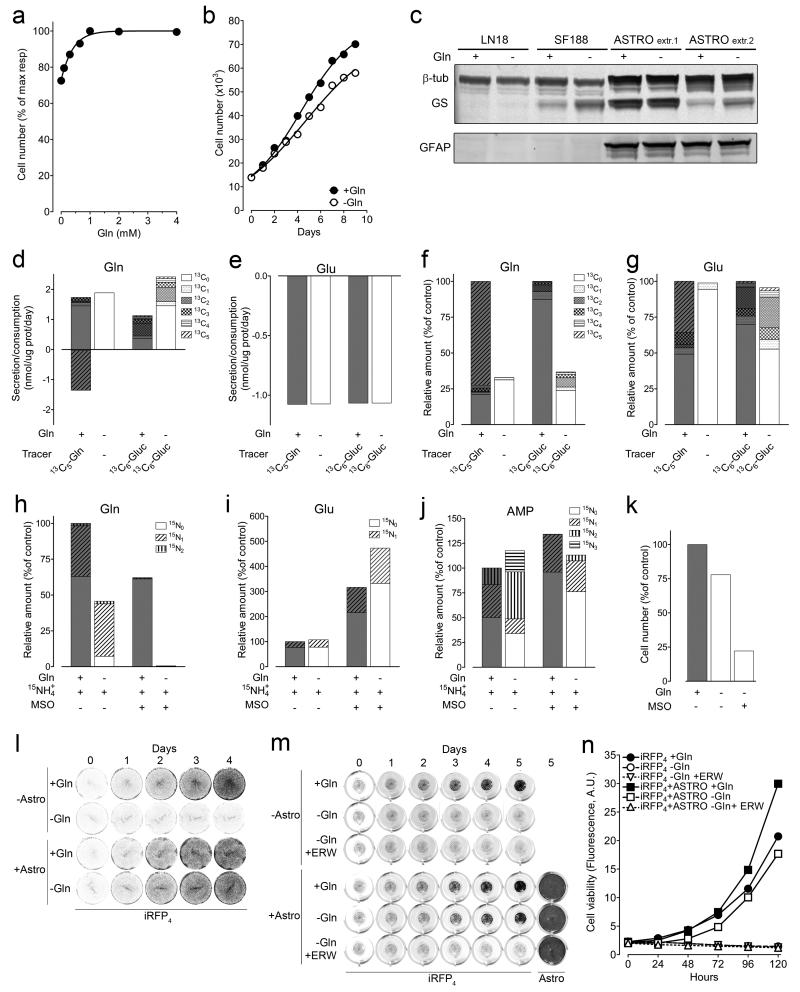
L-Glutamine (Gln) functions physiologically to balance the carbon and nitrogen requirements of tissues. It has been proposed that in cancer cells undergoing aerobic glycolysis, accelerated anabolism is sustained by Gln-derived carbons, which replenish the tricarboxylic acid (TCA) cycle (anaplerosis). However, it is shown here that in glioblastoma (GBM) cells, almost half of the Gln-derived glutamate (Glu) is secreted and does not enter the TCA cycle, and that inhibiting glutaminolysis does not affect cell proliferation. Moreover, Gln-starved cells are not rescued by TCA cycle replenishment. Instead, the conversion of Glu to Gln by glutamine synthetase (GS; cataplerosis) confers Gln prototrophy, and fuels de novo purine biosynthesis. In both orthotopic GBM models and in patients, (13)C-glucose tracing showed that GS produces Gln from TCA-cycle-derived carbons. Finally, the Gln required for the growth of GBM tumours is contributed only marginally by the circulation, and is mainly either autonomously synthesized by GS-positive glioma cells, or supplied by astrocytes.
Figures
Comment in
-
Rethinking glutamine addiction.Nat Cell Biol. 2015 Dec;17(12):1515-7. doi: 10.1038/ncb3278. Nat Cell Biol. 2015. PMID: 26612572
References
-
- Moreadith RW, Lehninger AL. The pathways of glutamate and glutamine oxidation by tumor cell mitochondria. Role of mitochondrial NAD(P)+-dependent malic enzyme. The Journal of biological chemistry. 1984;259:6215–6221. - PubMed
-
- Tardito S, et al. L-Asparaginase and inhibitors of glutamine synthetase disclose glutamine addiction of beta-catenin-mutated human hepatocellular carcinoma cells. Current cancer drug targets. 2011;11:929–943. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
