

Type I interferon restricts type 2 immunopathology through the regulation of group 2 innate lymphoid cells
- PMID: 26595887
- PMCID: PMC9135352
- DOI: 10.1038/ni.3308
Type I interferon restricts type 2 immunopathology through the regulation of group 2 innate lymphoid cells
Abstract
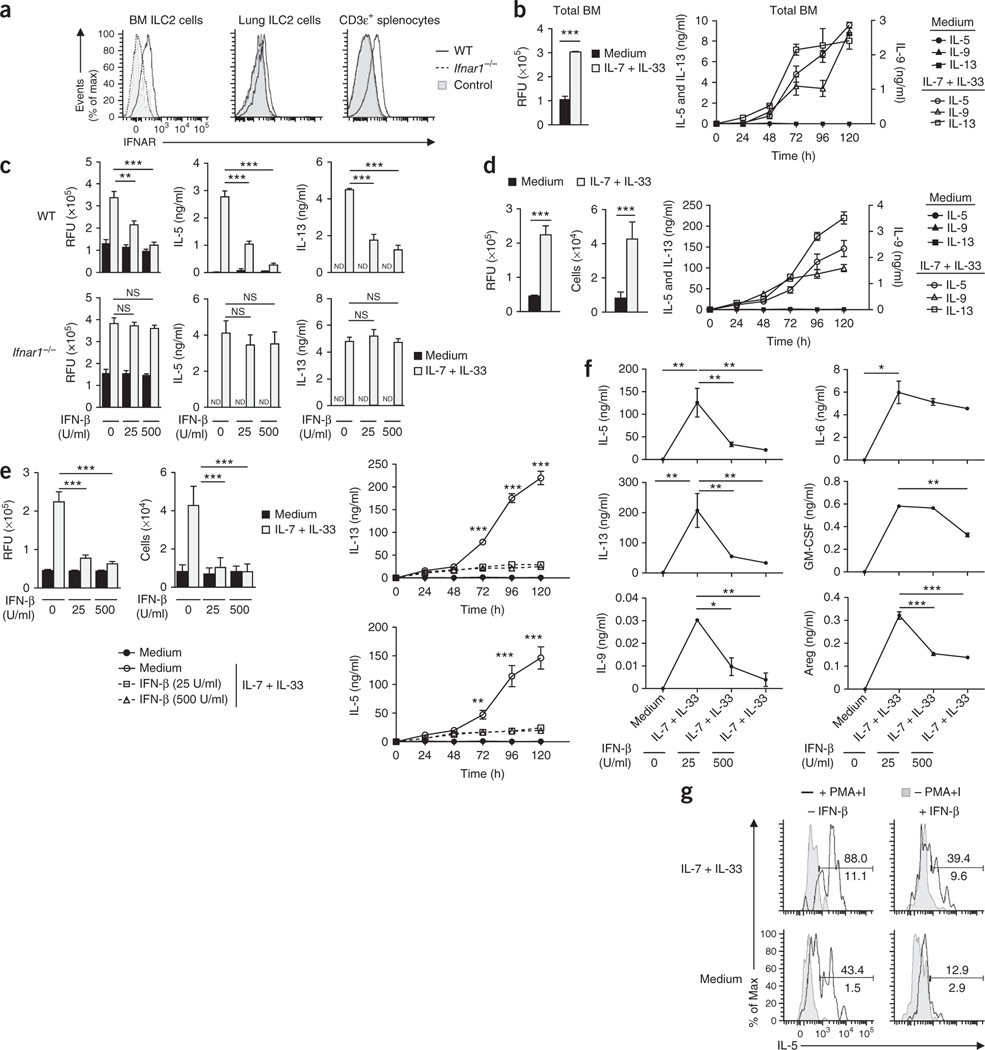
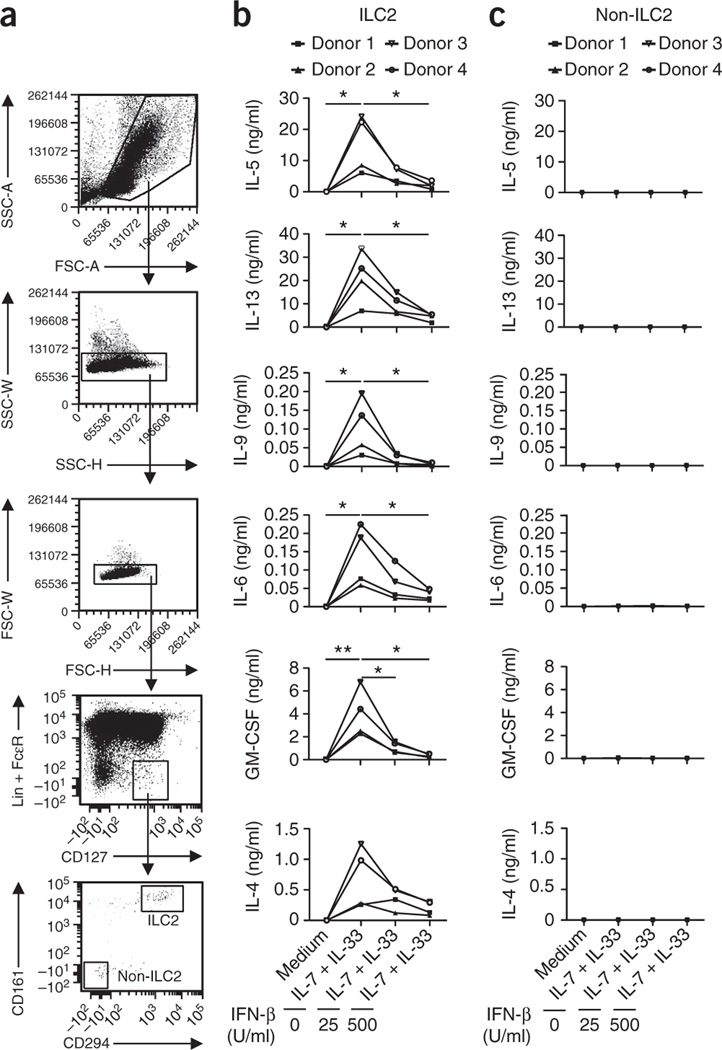
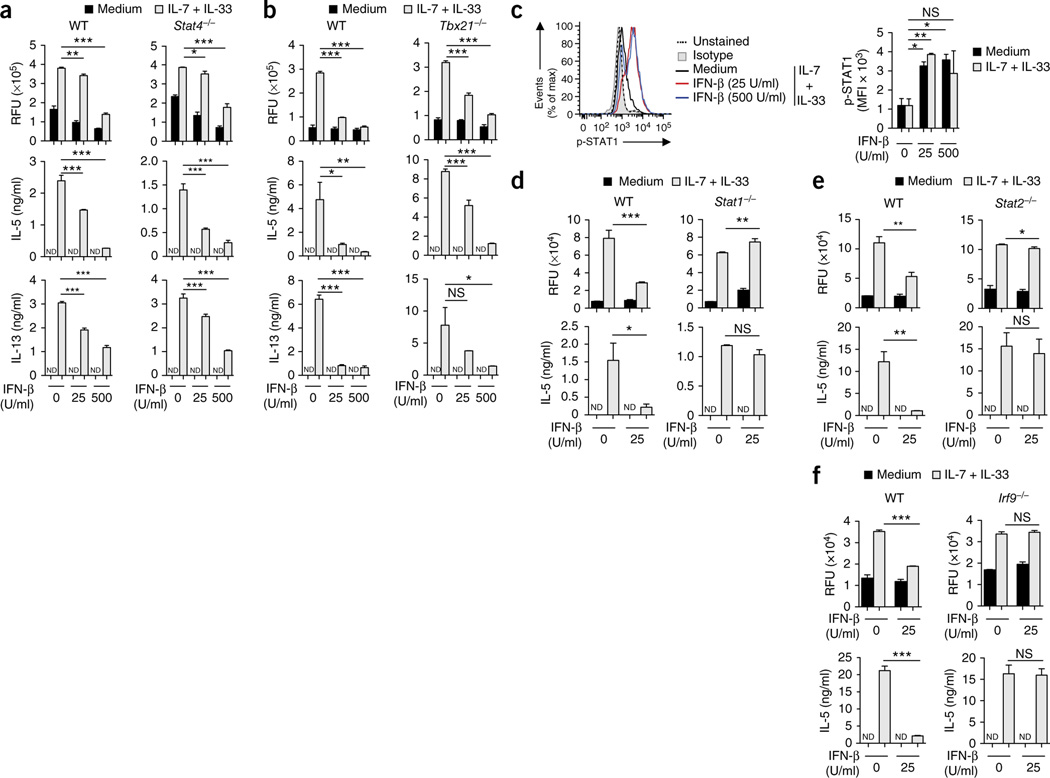
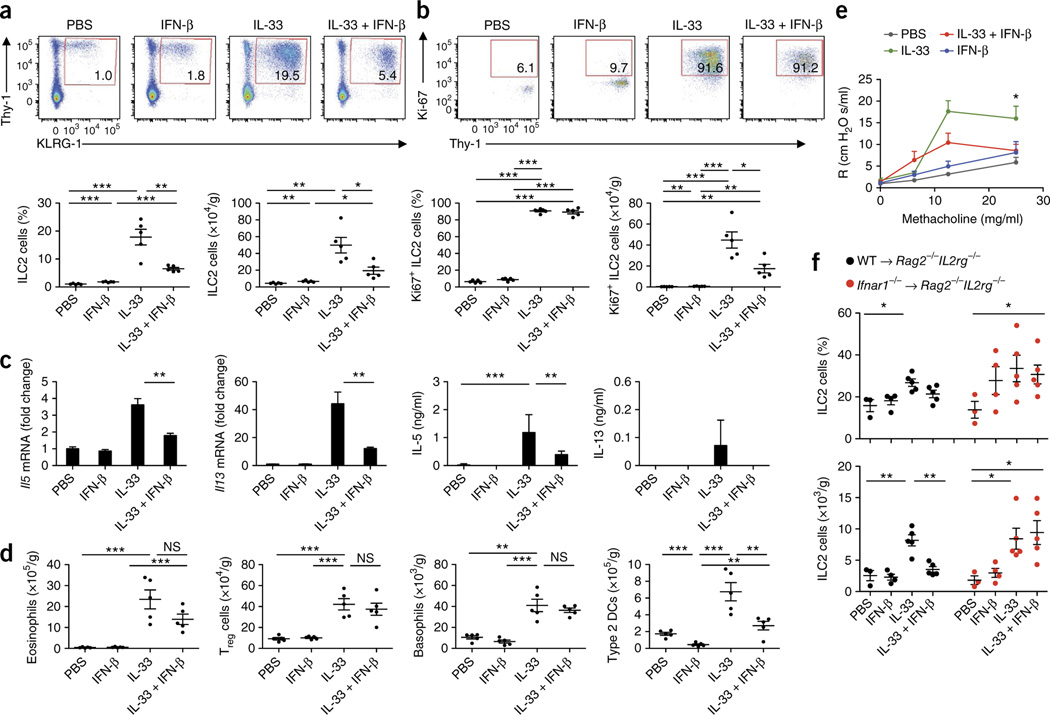
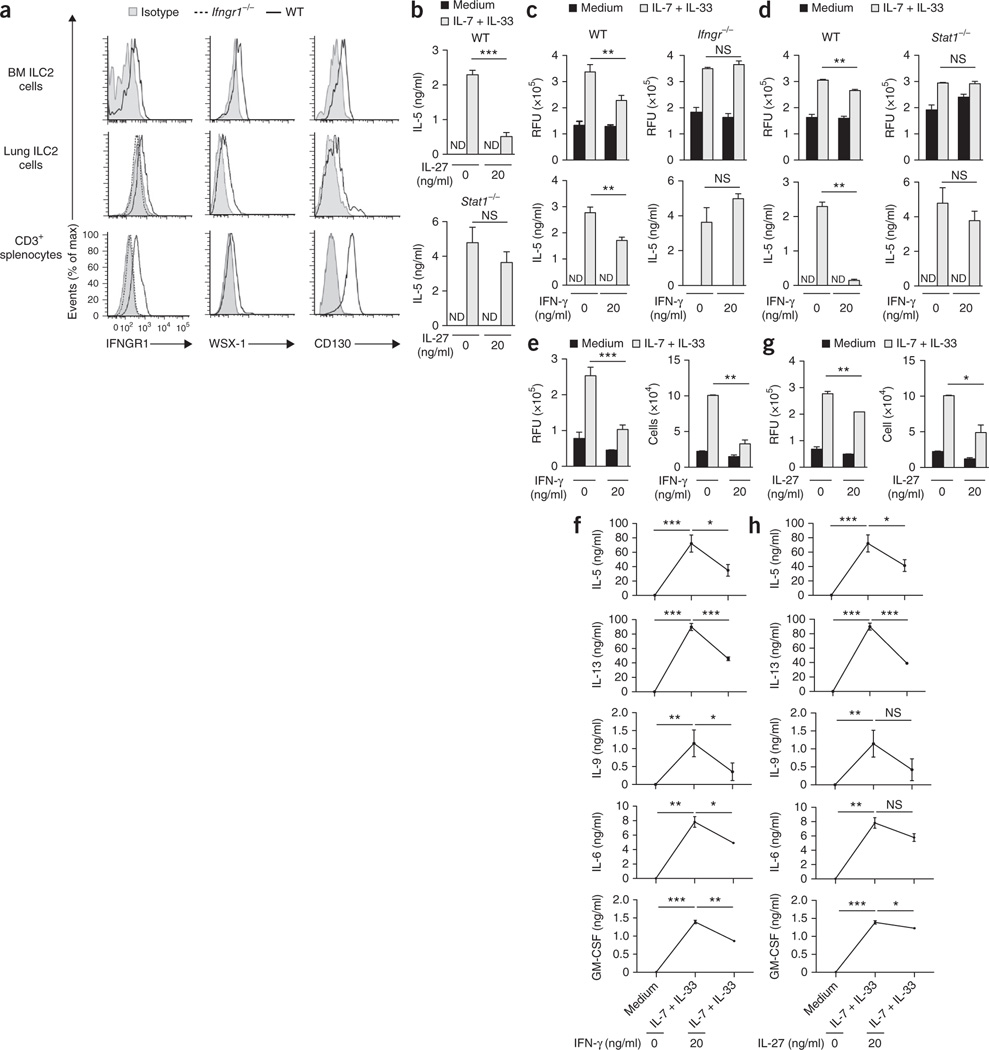
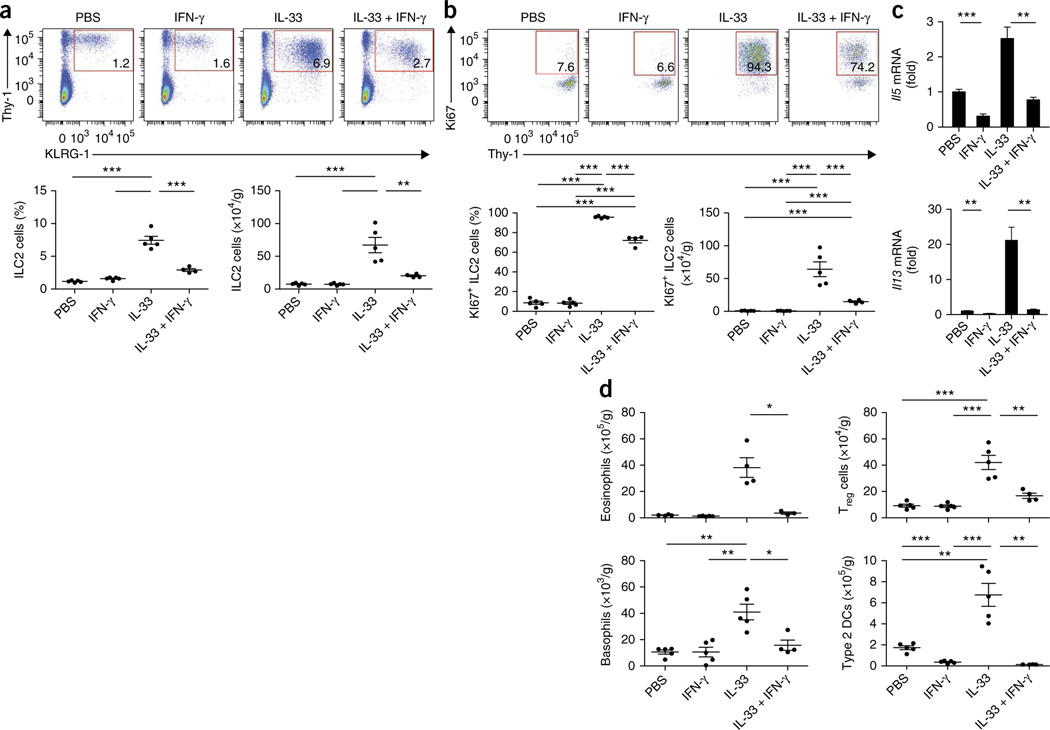
Viral respiratory tract infections are the main causative agents of the onset of infection-induced asthma and asthma exacerbations that remain mechanistically unexplained. Here we found that deficiency in signaling via type I interferon receptor led to deregulated activation of group 2 innate lymphoid cells (ILC2 cells) and infection-associated type 2 immunopathology. Type I interferons directly and negatively regulated mouse and human ILC2 cells in a manner dependent on the transcriptional activator ISGF3 that led to altered cytokine production, cell proliferation and increased cell death. In addition, interferon-γ (IFN-γ) and interleukin 27 (IL-27) altered ILC2 function dependent on the transcription factor STAT1. These results demonstrate that type I and type II interferons, together with IL-27, regulate ILC2 cells to restrict type 2 immunopathology.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
