

Complementarity and redundancy of IL-22-producing innate lymphoid cells
- PMID: 26595889
- PMCID: PMC4720992
- DOI: 10.1038/ni.3332
Complementarity and redundancy of IL-22-producing innate lymphoid cells
Abstract
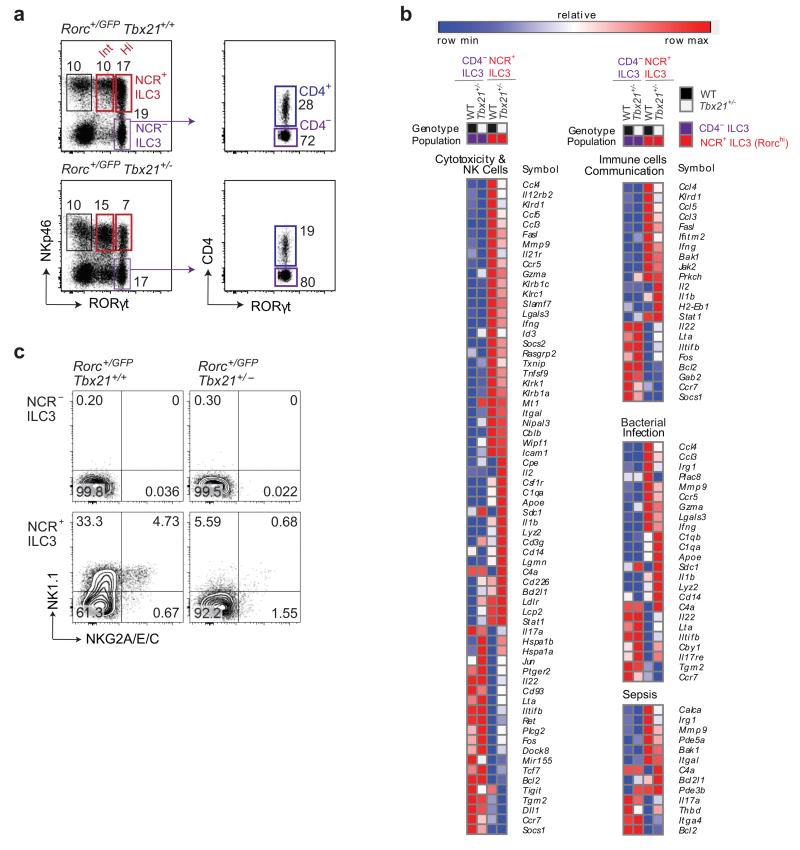
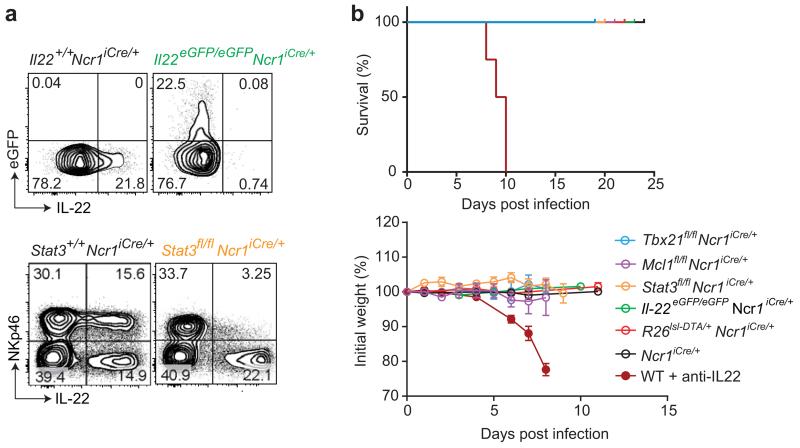
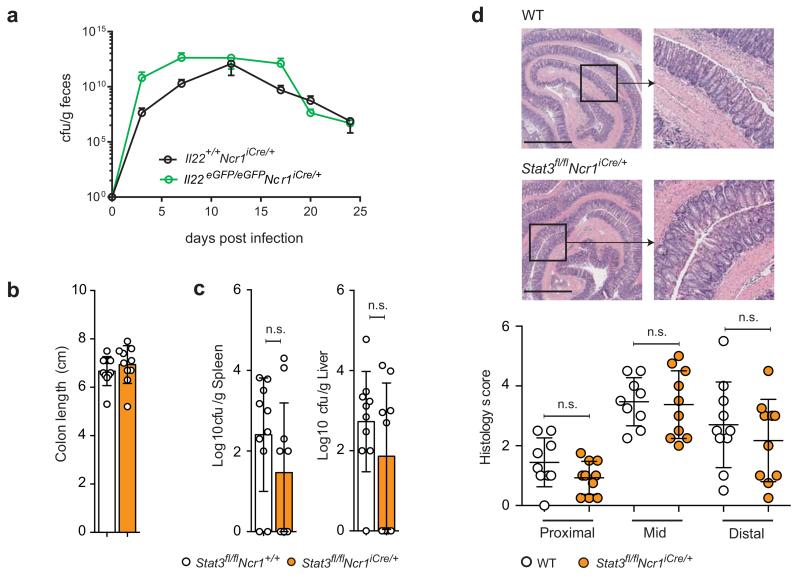
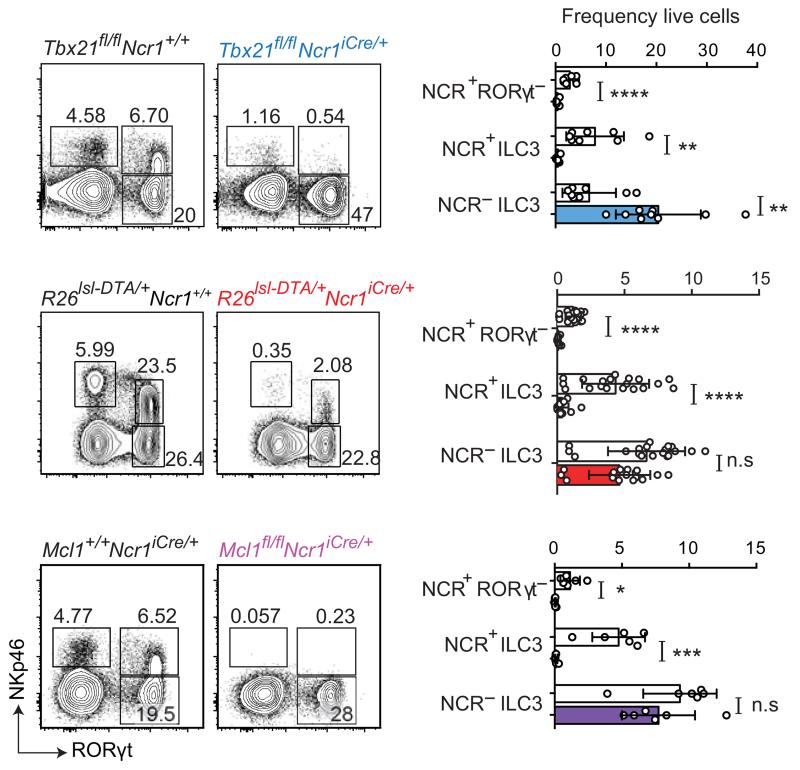
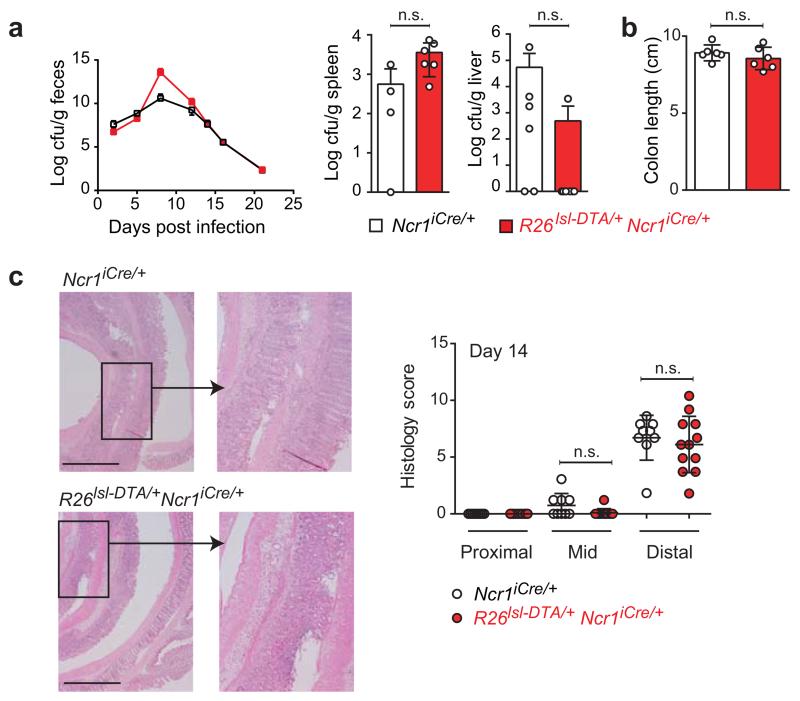
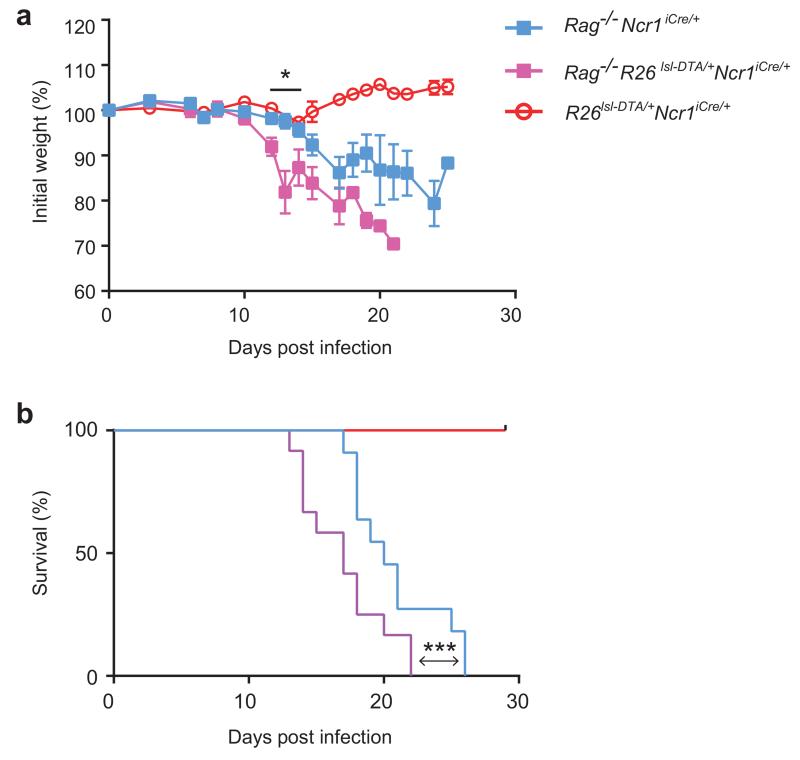
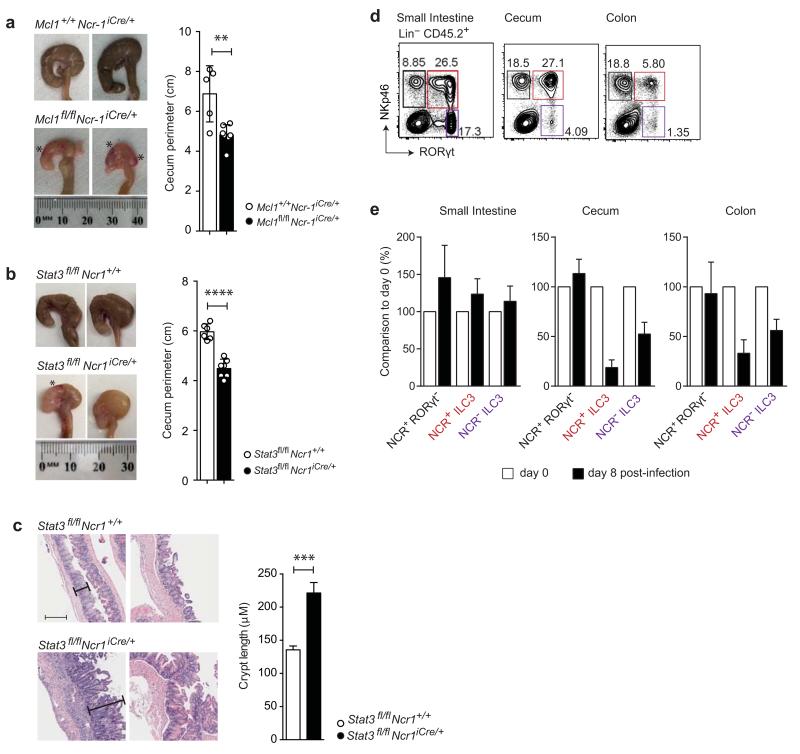
Intestinal T cells and group 3 innate lymphoid cells (ILC3 cells) control the composition of the microbiota and gut immune responses. Within the gut, ILC3 subsets coexist that either express or lack the natural cytoxicity receptor (NCR) NKp46. We identified here the transcriptional signature associated with the transcription factor T-bet-dependent differentiation of NCR(-) ILC3 cells into NCR(+) ILC3 cells. Contrary to the prevailing view, we found by conditional deletion of the key ILC3 genes Stat3, Il22, Tbx21 and Mcl1 that NCR(+) ILC3 cells were redundant for the control of mouse colonic infection with Citrobacter rodentium in the presence of T cells. However, NCR(+) ILC3 cells were essential for cecal homeostasis. Our data show that interplay between intestinal ILC3 cells and adaptive lymphocytes results in robust complementary failsafe mechanisms that ensure gut homeostasis.
Figures
References
-
- Spits H, Artis D, Colonna M, Diefenbach A, Di Santo JP, Eberl G, et al. Innate lymphoid cells--a proposal for uniform nomenclature. Nat Rev Immunol. 2013;13:145–149. - PubMed
-
- McKenzie AN, Spits H, Eberl G. Innate lymphoid cells in inflammation and immunity. Immunity. 2014;41:366–374. - PubMed
-
- Montaldo E, Juelke K, Romagnani C. Group 3 innate lymphoid cells (ILC3s): Origin, differentiation, and plasticity in humans and mice. Eur J Immunol. 2015;45:2171–2182. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
