

Single-molecule enzymology of steroid transforming enzymes: Transient kinetic studies and what they tell us
- PMID: 26596239
- PMCID: PMC4842339
- DOI: 10.1016/j.jsbmb.2015.10.016
Single-molecule enzymology of steroid transforming enzymes: Transient kinetic studies and what they tell us
Abstract
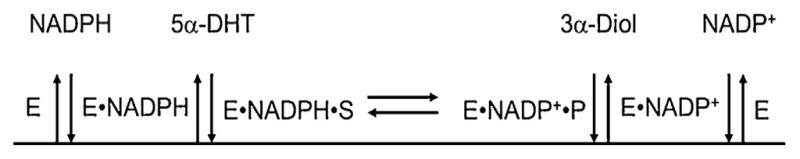
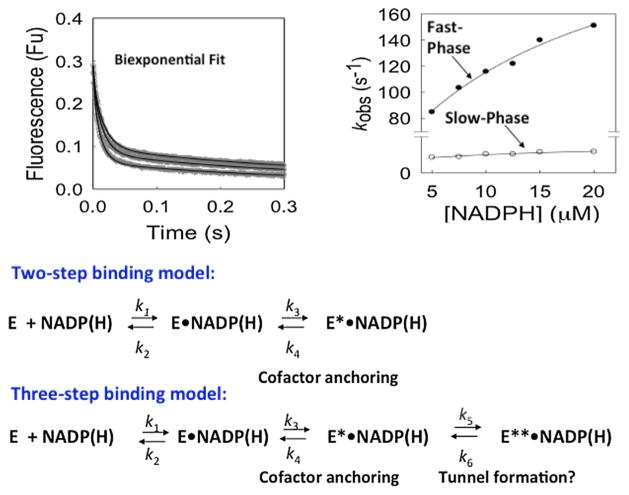
Structure-function studies on steroid transforming enzymes often use site-directed mutagenesis to inform mechanisms of catalysis and effects on steroid binding, and data are reported in terms of changes in steady state kinetic parameters kcat, Km and kcat/Km. However, this dissection of function is limited since kcat is governed by the rate-determining step and Km is a complex macroscopic kinetic constant. Often site-directed mutagenesis can lead to a change in the rate-determining step which cannot be revealed by just reporting a decrease in kcat alone. These issues are made more complex when it is considered that many steroid transforming enzymes have more than one substrate and product. We present the case for using transient-kinetics performed with stopped-flow spectrometry to assign rate constants to discrete steps in these multi-substrate reactions and their use to interpret enzyme mechanism and the effects of disease and engineered mutations. We demonstrate that fluorescence kinetic transients can be used to measure ligand binding that may be accompanied by isomerization steps, revealing the existence of new enzyme intermediates. We also demonstrate that single-turnover reactions can provide a klim for the chemical step and Ks for steroid-substrate binding and that when coupled with kinetic isotope effect measurements can provide information on transition state intermediates. We also demonstrate how multiple turnover experiments can provide evidence for either "burst-phase" kinetics, which can reveal a slow product release step, or linear-phase kinetics, in which the chemical step can be rate-determining. With these assignments it becomes more straightforward to analyze the effects of mutations. We use examples from the hydroxysteroid dehydrogenases (AKR1Cs) and human steroid 5β-reductase (AKR1D1) to illustrate the utility of the approach, which are members of the aldo-keto reductase (AKR) superfamily.
Keywords: Aldo-keto reductase; Hydroxysteroid dehydrogenase; Kinetic-isotope effect; Steroid double-bond reductases; Stopped-flow; Transient-kientics.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures





References
-
- Michealis L, Menten ML. Kinetik der Invertinwirkung. Biochemishches Zietschrift. 1913;49:333–369.
-
- Boehmer AL, Brinkmann AO, Sandkuijl LA, Halley DJ, Niermeijer MF, Andersson S, de Jong FH, Kayserili H, de Vroede MA, Otten BJ, Rouwé CW, Mendonça BB, Rodrigues C, Bode HH, de Ruiter PE, de Waal Delemarre-van HA, Drop SL. 17β-hydroxysteroid dehydrogenase-3 deficiency: diagnosis, phenotypic variability, population genetics, and worldwide distribution of ancient and de novo mutations. J Clin Endocrinol Metab. 1999;84:4713–4721. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
