

Proteomic Analysis of Dynein-Interacting Proteins in Amyotrophic Lateral Sclerosis Synaptosomes Reveals Alterations in the RNA-Binding Protein Staufen1
- PMID: 26598648
- PMCID: PMC4739670
- DOI: 10.1074/mcp.M115.049965
Proteomic Analysis of Dynein-Interacting Proteins in Amyotrophic Lateral Sclerosis Synaptosomes Reveals Alterations in the RNA-Binding Protein Staufen1
Abstract
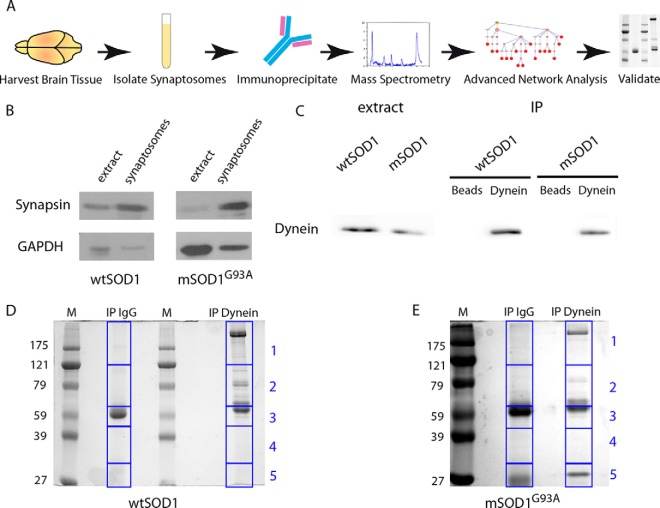
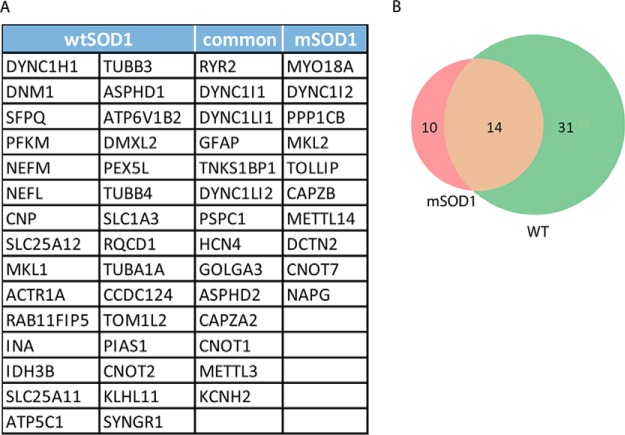
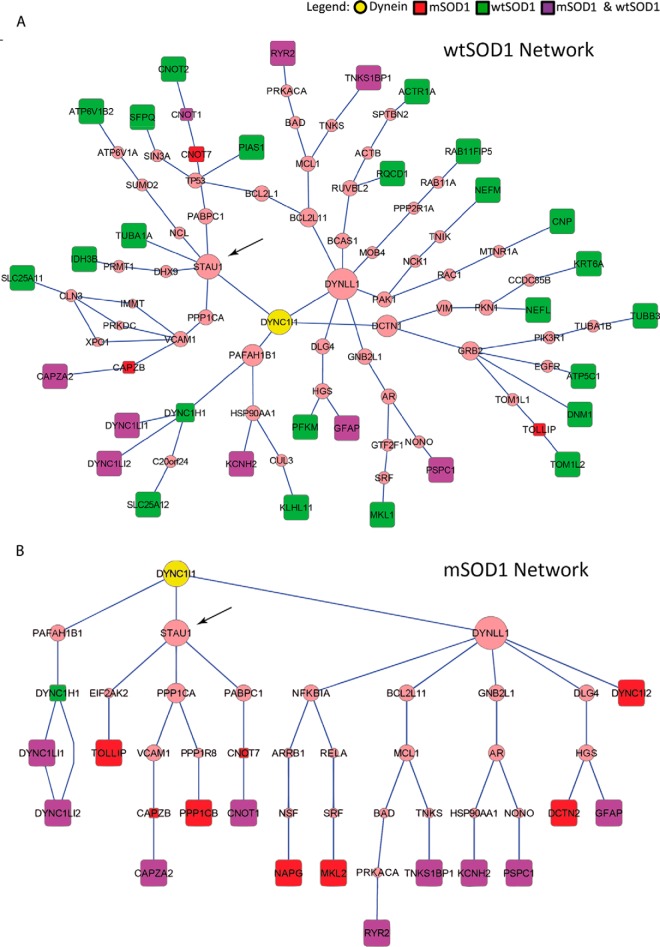
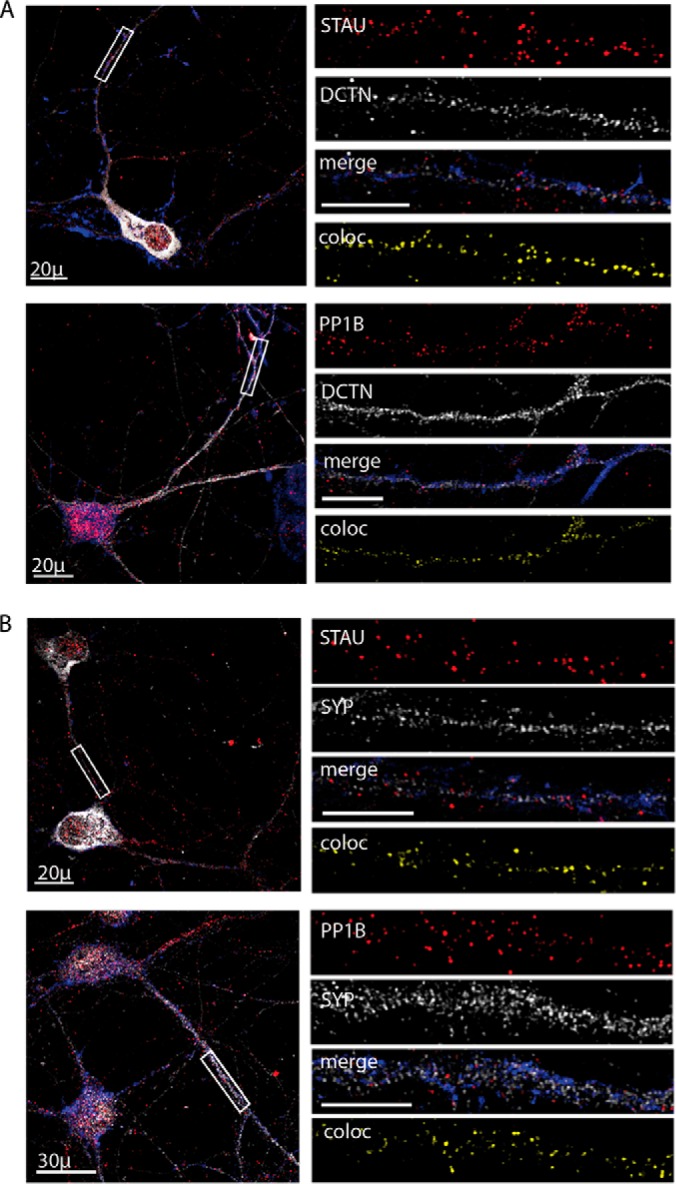
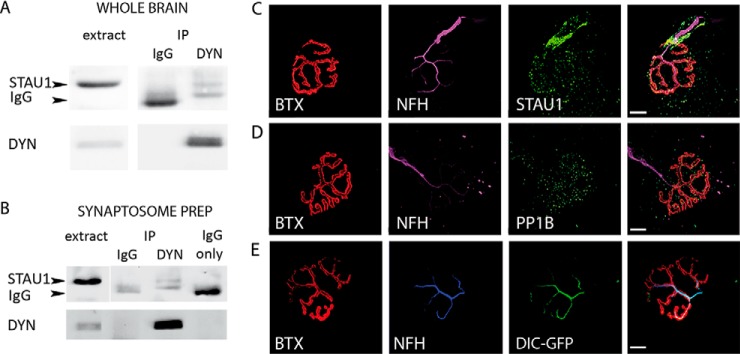
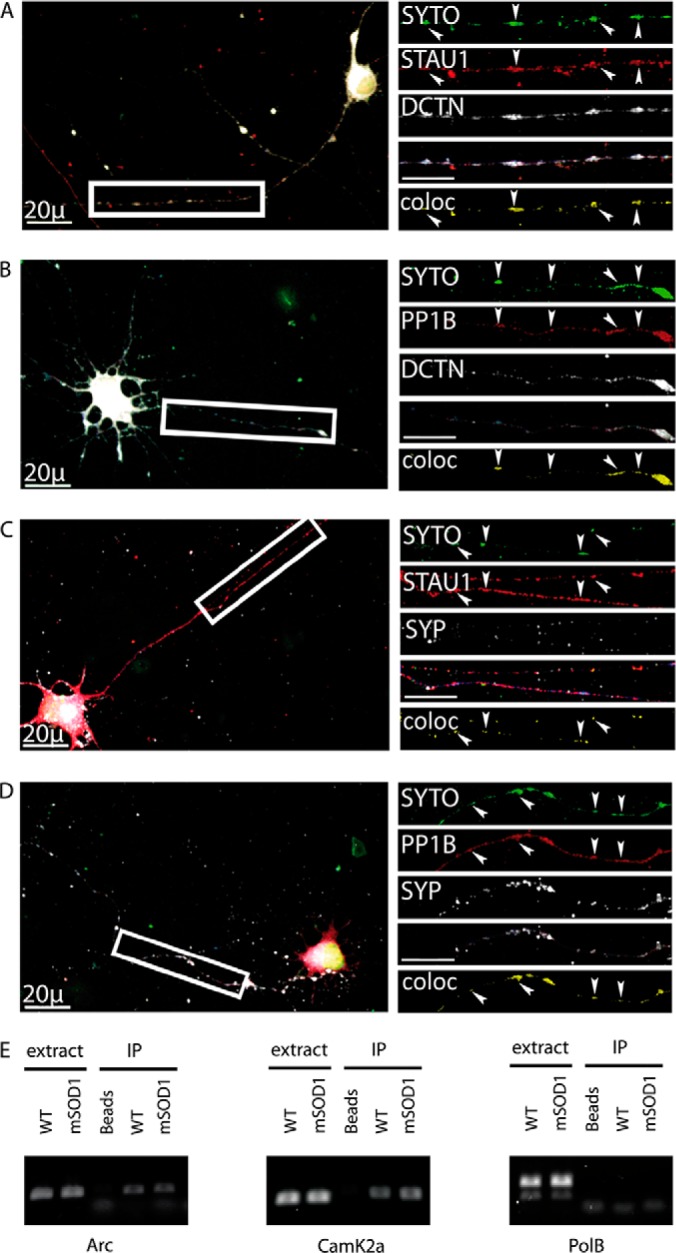
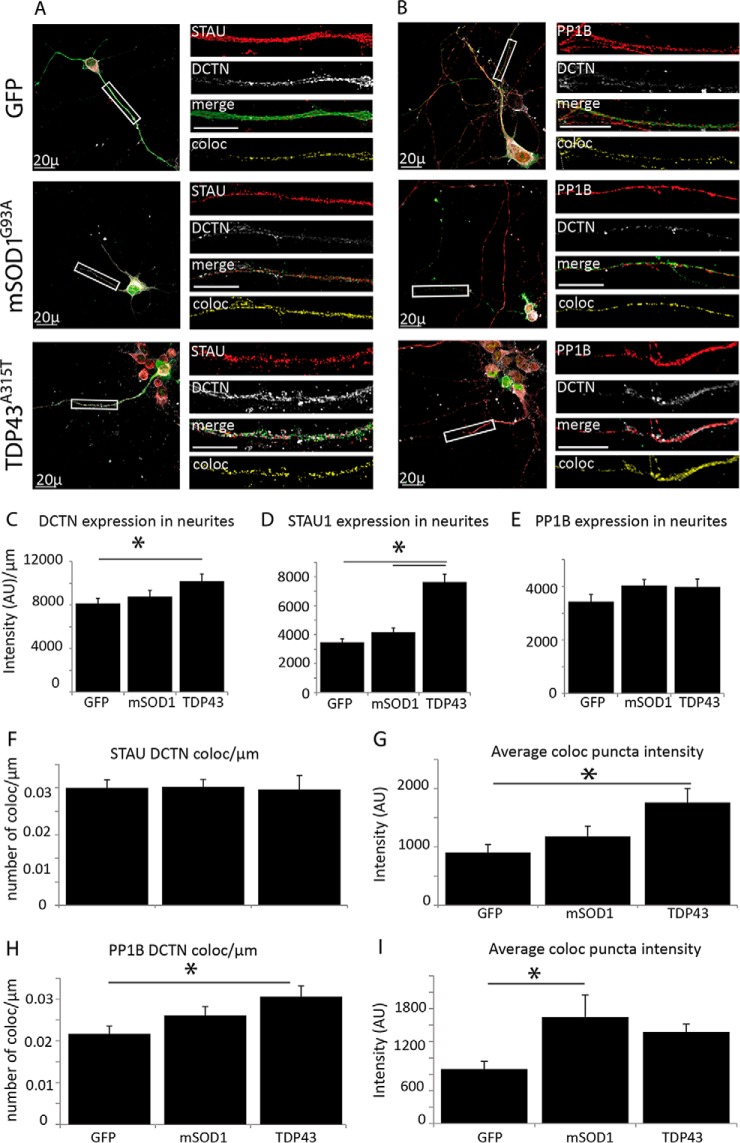
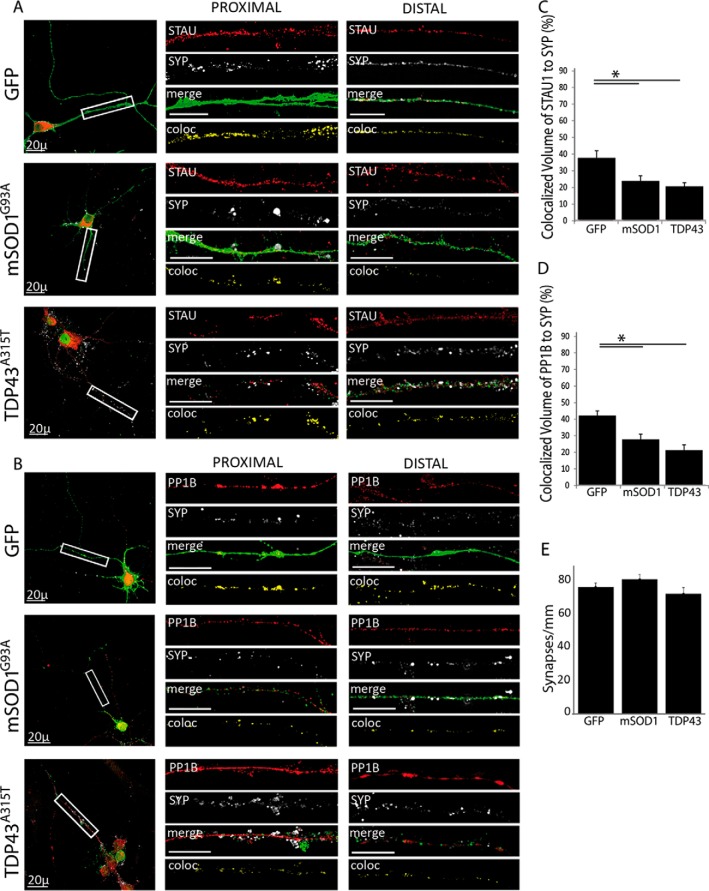
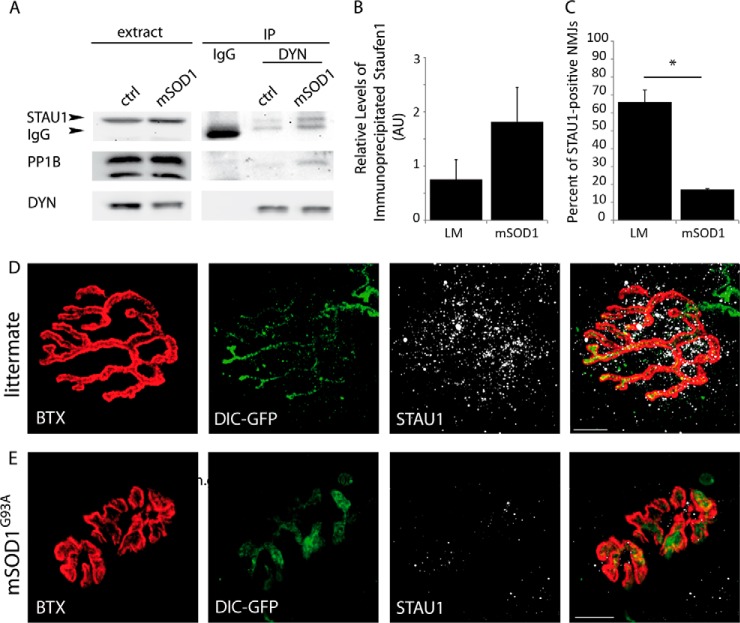
Synapse disruption takes place in many neurodegenerative diseases, including amyotrophic lateral sclerosis (ALS). However, the mechanistic understanding of this process is still limited. We set out to study a possible role for dynein in synapse integrity. Cytoplasmic dynein is a multisubunit intracellular molecule responsible for diverse cellular functions, including long-distance transport of vesicles, organelles, and signaling factors toward the cell center. A less well-characterized role dynein may play is the spatial clustering and anchoring of various factors including mRNAs in distinct cellular domains such as the neuronal synapse. Here, in order to gain insight into dynein functions in synapse integrity and disruption, we performed a screen for novel dynein interactors at the synapse. Dynein immunoprecipitation from synaptic fractions of the ALS model mSOD1(G93A) and wild-type controls, followed by mass spectrometry analysis on synaptic fractions of the ALS model mSOD1(G93A) and wild-type controls, was performed. Using advanced network analysis, we identified Staufen1, an RNA-binding protein required for the transport and localization of neuronal RNAs, as a major mediator of dynein interactions via its interaction with protein phosphatase 1-beta (PP1B). Both in vitro and in vivo validation assays demonstrate the interactions of Staufen1 and PP1B with dynein, and their colocalization with synaptic markers was altered as a result of two separate ALS-linked mutations: mSOD1(G93A) and TDP43(A315T). Taken together, we suggest a model in which dynein's interaction with Staufen1 regulates mRNA localization along the axon and the synapses, and alterations in this process may correlate with synapse disruption and ALS toxicity.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- De Vos K. J., Grierson A. J., Ackerley S., and Miller C. C. (2008) Role of axonal transport in neurodegenerative diseases. Annu. Rev. Neurosci. 31, 151–173 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
