

A flexible genetic toolkit for arthropod neurogenesis
- PMID: 26598727
- PMCID: PMC4685583
- DOI: 10.1098/rstb.2015.0044
A flexible genetic toolkit for arthropod neurogenesis
Abstract
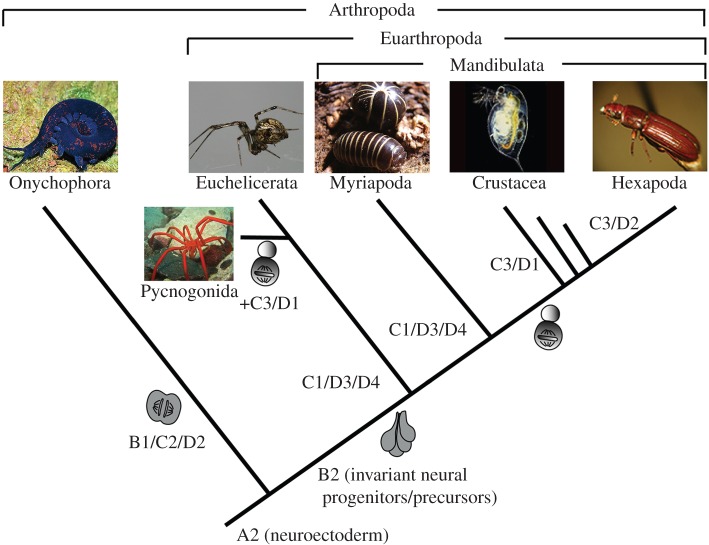
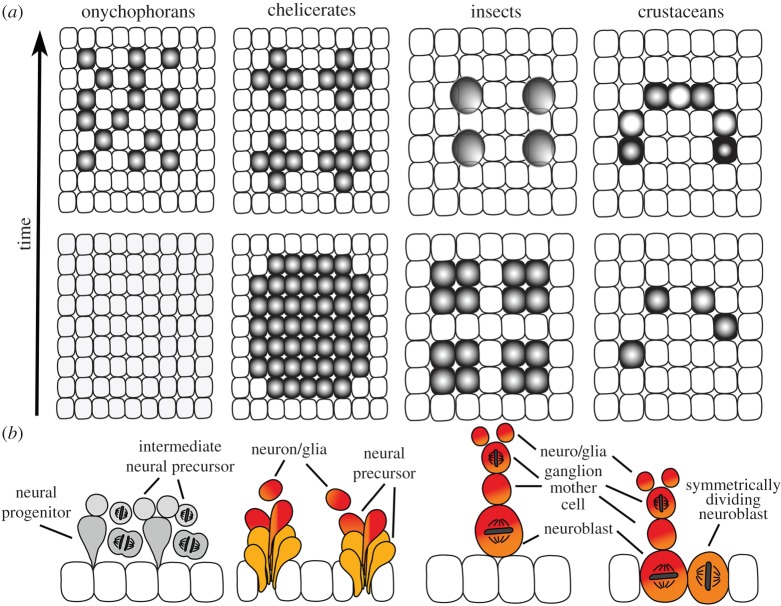
Arthropods show considerable variations in early neurogenesis. This includes the pattern of specification, division and movement of neural precursors and progenitors. In all metazoans with nervous systems, including arthropods, conserved genes regulate neurogenesis, which raises the question of how the various morphological mechanisms have emerged and how the same genetic toolkit might generate different morphological outcomes. Here I address this question by comparing neurogenesis across arthropods and show how variations in the regulation and function of the neural genes might explain this phenomenon and how they might have facilitated the evolution of the diverse morphological mechanisms of neurogenesis.
Keywords: Notch signalling; asymmetric division; neural precursors; neural progenitors; neurogenic potential; patterning genes.
© 2015 The Author(s).
Figures

References
-
- Storch V, Welsch U. 2004. Systematische zoologie. Berlin, Germany: Spektrum akademischer Verlag.
-
- Rota-Stabelli O, Campbell L, Brinkmann H, Edgecombe GD, Longhorn SJ, Peterson KJ, Pisani D, Philippe H, Telford MJ. 2011. A congruent solution to arthropod phylogeny: phylogenomics, microRNAs and morphology support monophyletic Mandibulata. Proc. R. Soc. B 278, 298–306. (10.1098/rspb.2010.0590) - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
