

Mutually Exclusive Roles of SHARPIN in Integrin Inactivation and NF-κB Signaling
- PMID: 26600301
- PMCID: PMC4658161
- DOI: 10.1371/journal.pone.0143423
Mutually Exclusive Roles of SHARPIN in Integrin Inactivation and NF-κB Signaling
Abstract
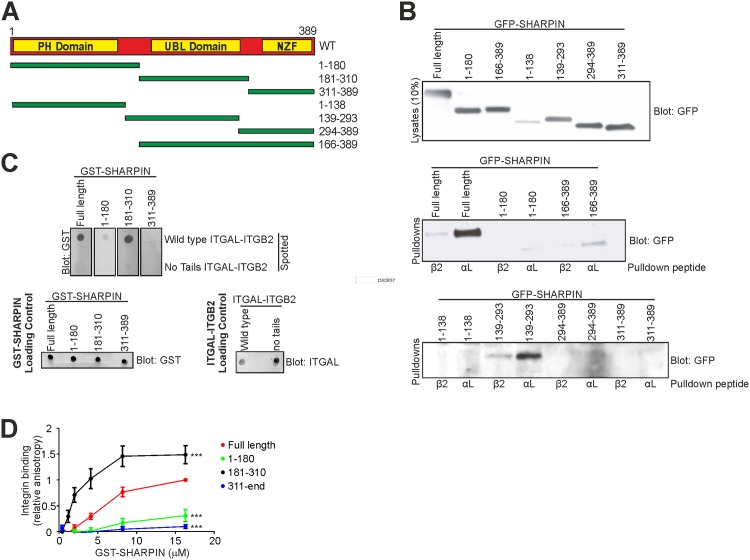
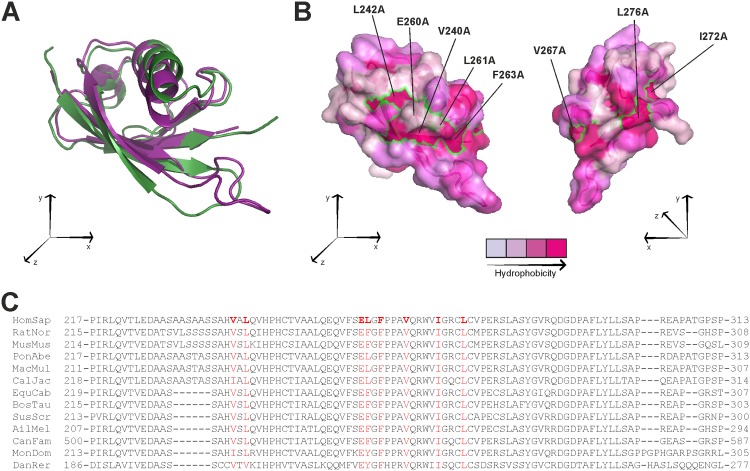
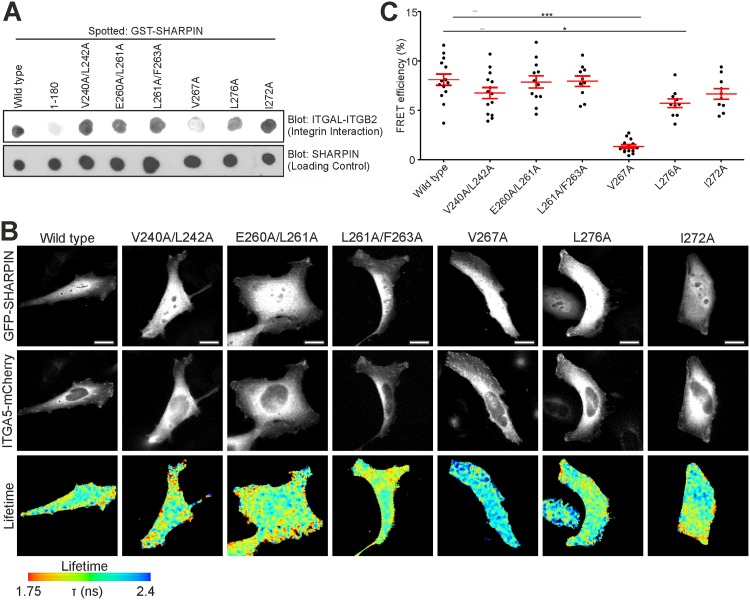
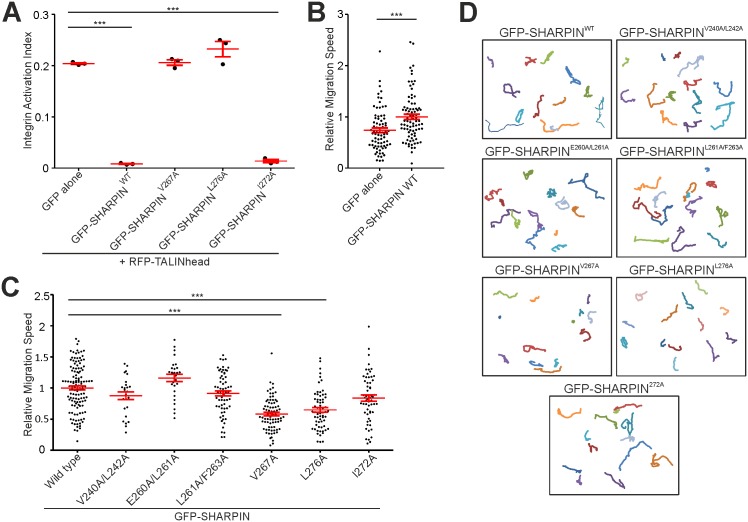
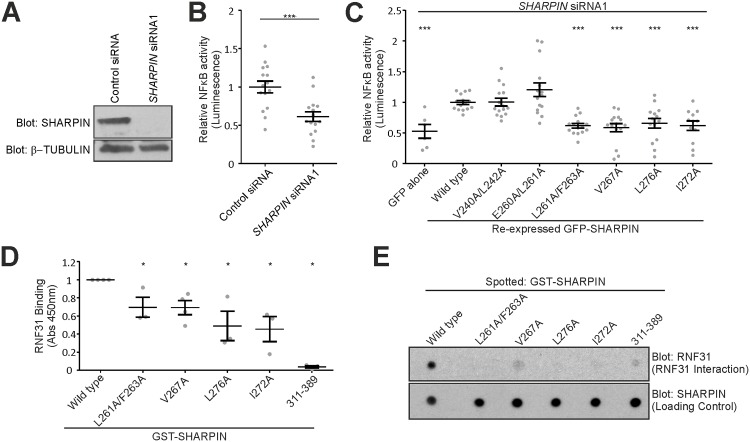
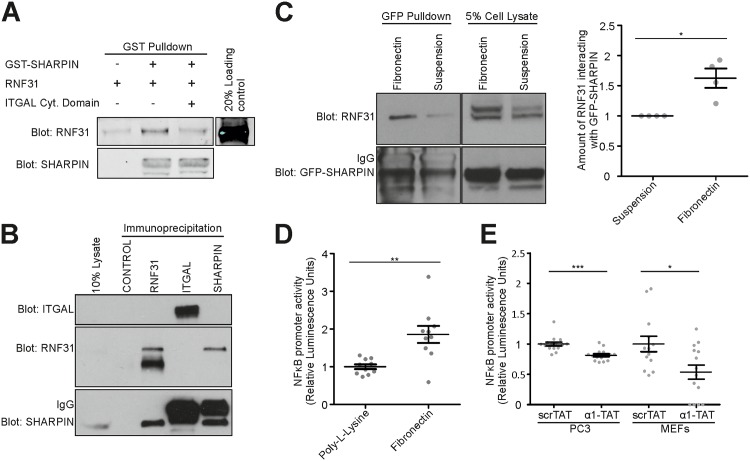
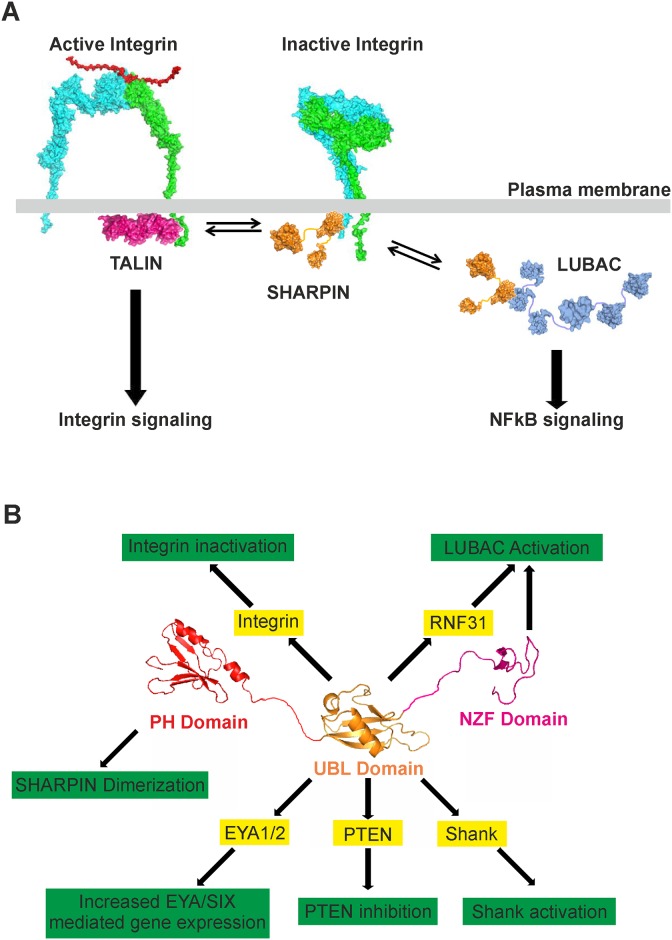
SHANK-associated RH domain interactor (SHARPIN) inhibits integrins through interaction with the integrin α-subunit. In addition, SHARPIN enhances nuclear factor-kappaB (NF-κB) activity as a component of the linear ubiquitin chain assembly complex (LUBAC). However, it is currently unclear how regulation of these seemingly different roles is coordinated. Here, we show that SHARPIN binds integrin and LUBAC in a mutually exclusive manner. We map the integrin binding site on SHARPIN to the ubiquitin-like (UBL) domain, the same domain implicated in SHARPIN interaction with LUBAC component RNF31 (ring finger protein 31), and identify two SHARPIN residues (V267, L276) required for both integrin and RNF31 regulation. Accordingly, the integrin α-tail is capable of competing with RNF31 for SHARPIN binding in vitro. Importantly, the full SHARPIN RNF31-binding site contains residues (F263A/I272A) that are dispensable for SHARPIN-integrin interaction. Importantly, disrupting SHARPIN interaction with integrin or RNF31 abolishes SHARPIN-mediated regulation of integrin or NF-κB activity, respectively. Altogether these data suggest that the roles of SHARPIN in inhibiting integrin activity and supporting linear ubiquitination are (molecularly) distinct.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
