

Locomotor play drives motor skill acquisition at the expense of growth: A life history trade-off
- PMID: 26601237
- PMCID: PMC4643810
- DOI: 10.1126/sciadv.1500451
Locomotor play drives motor skill acquisition at the expense of growth: A life history trade-off
Abstract
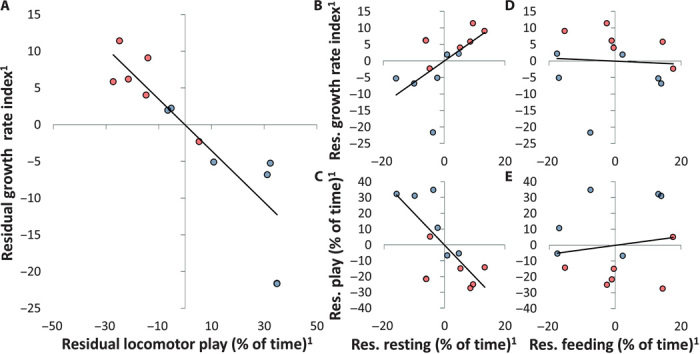
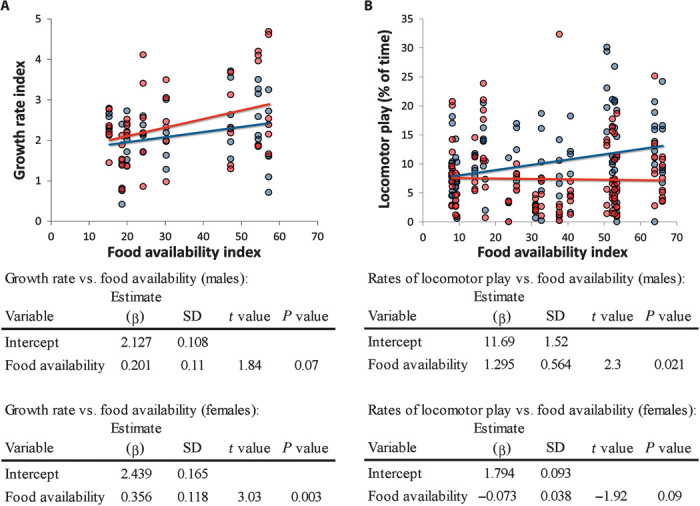
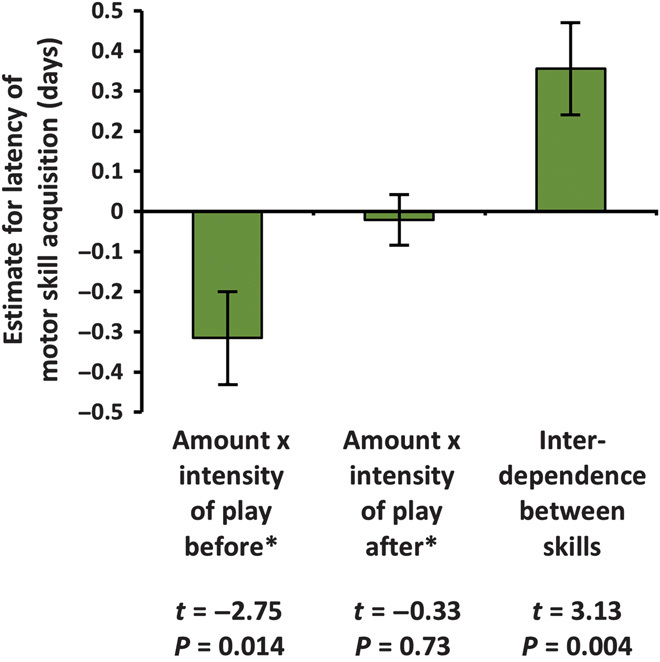
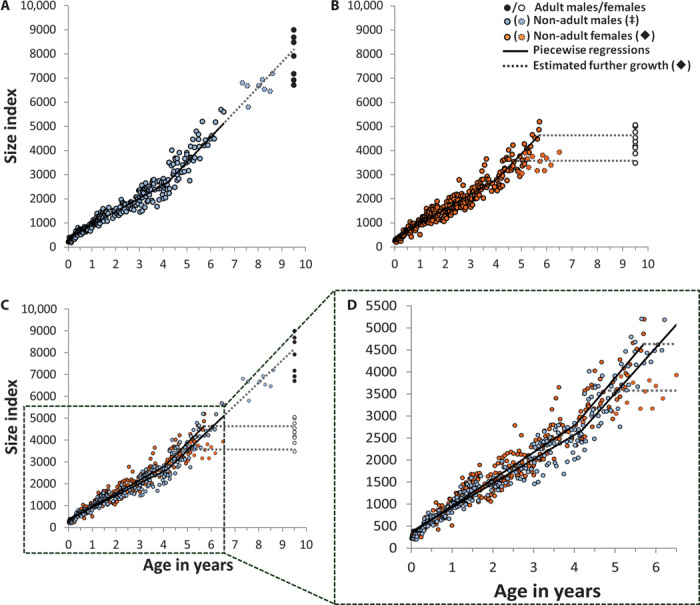
The developmental costs and benefits of early locomotor play are a puzzling topic in biology, psychology, and health sciences. Evolutionary theory predicts that energy-intensive behavior such as play can only evolve if there are considerable benefits. Prominent theories propose that locomotor play is (i) low cost, using surplus energy remaining after growth and maintenance, and (ii) beneficial because it trains motor skills. However, both theories are largely untested. Studying wild Assamese macaques, we combined behavioral observations of locomotor play and motor skill acquisition with quantitative measures of natural food availability and individual growth rates measured noninvasively via photogrammetry. Our results show that investments in locomotor play were indeed beneficial by accelerating motor skill acquisition but carried sizable costs in terms of reduced growth. Even under moderate natural energy restriction, investment in locomotor play accounted for up to 50% of variance in growth, which strongly contradicts the current theory that locomotor play only uses surplus energy remaining after growth and maintenance. Male immatures played more, acquired motor skills faster, and grew less than female immatures, leading to persisting size differences until the age of female maturity. Hence, depending on skill requirements, investment in play can take ontogenetic priority over physical development unconstrained by costs of play with consequences for life history, which strongly highlights the ontogenetic and evolutionary importance of play.
Keywords: Developmental origins of health and disease; human evolution; juvenile risk hypothesis; motor training hypothesis; phenotypic plasticity; resource allocation; surplus resource hypothesis.
Figures
References
-
- P. M. Kappeler, M. E. Pereira, Primate Life Histories and Socioecology (The Univ. of Chicago Press, Chicago, 2003).
-
- Pontzer H., Raichlen D. A., Gordon A. D., Schroepfer-Walker K. K., Hare B., O’Neill M. C., Muldoon K. M., Dunsworth H. M., Wood B. M., Isler K., Burkart J., Irwin M., Shumaker R. W., Lonsdorf E. V., Primate energy expenditure and life history. Proc. Natl. Acad. Sci. U.S.A. 111, 1433–1437 (2014). - PMC - PubMed
-
- S. C. Stearns, The Evolution of Life Histories (Oxford Univ. Press, Oxford, 1992).
-
- Brown J. H., Gillooly J. F., Allen A. P., Savage V. M., West G. B., Toward a metabolic theory of ecology. Ecology 85, 1771–1789 (2004).
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
