

Cis-regulatory mechanisms governing stem and progenitor cell transitions
- PMID: 26601269
- PMCID: PMC4643771
- DOI: 10.1126/sciadv.1500503
Cis-regulatory mechanisms governing stem and progenitor cell transitions
Abstract
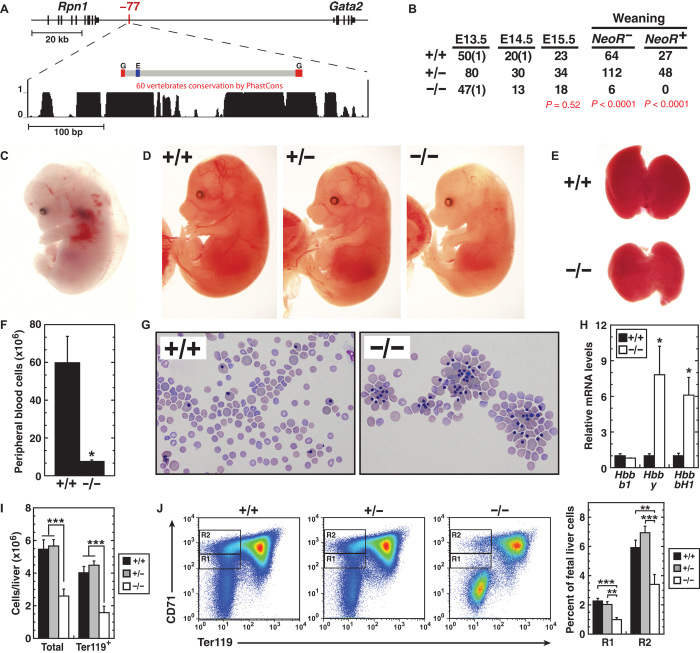
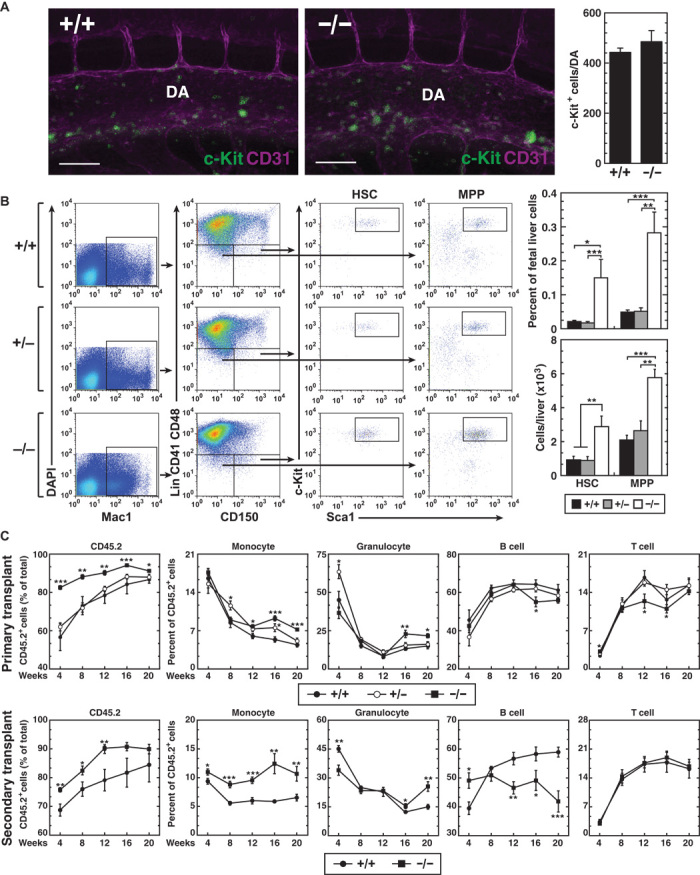
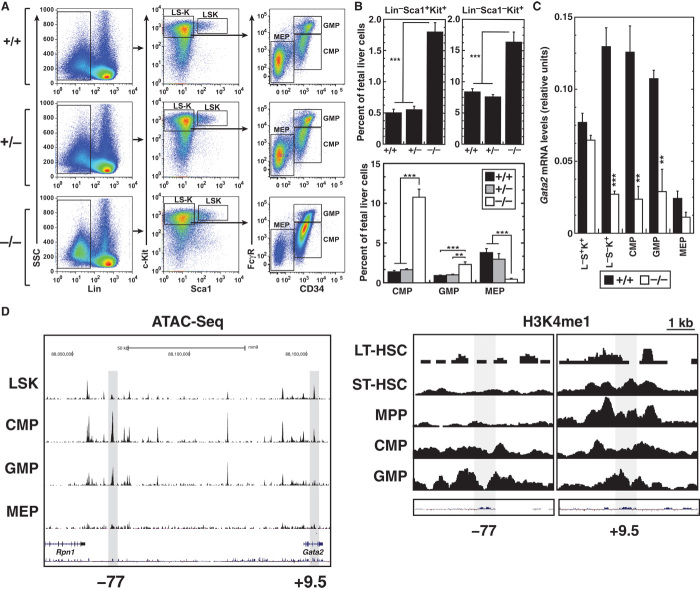
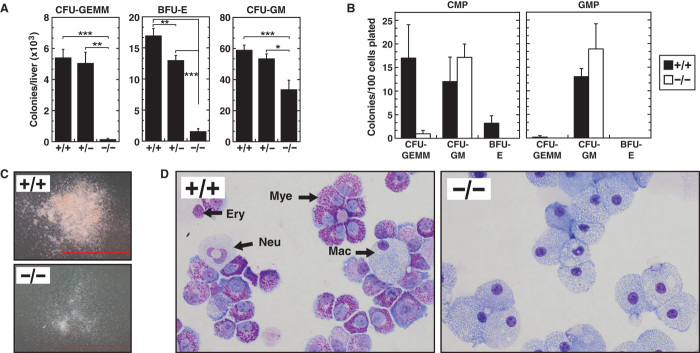
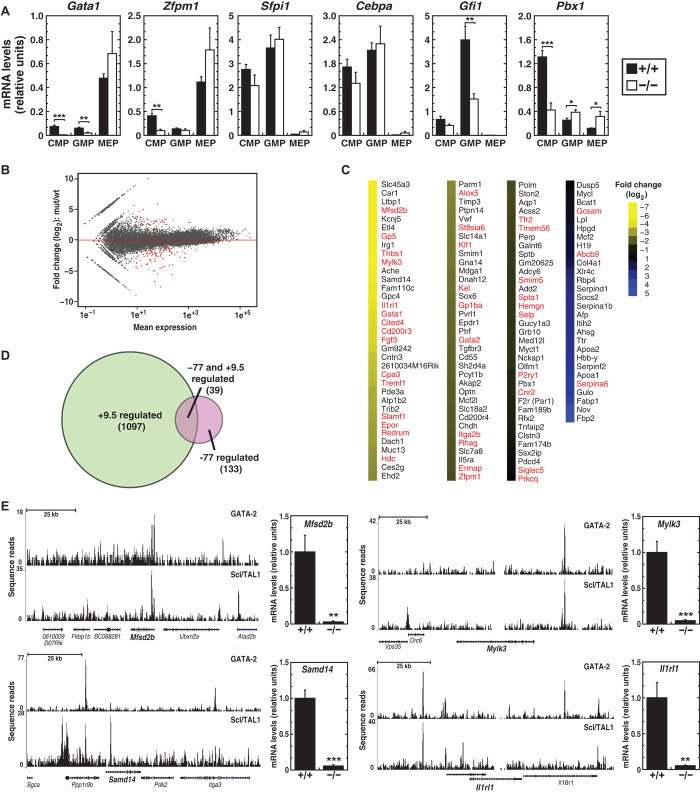
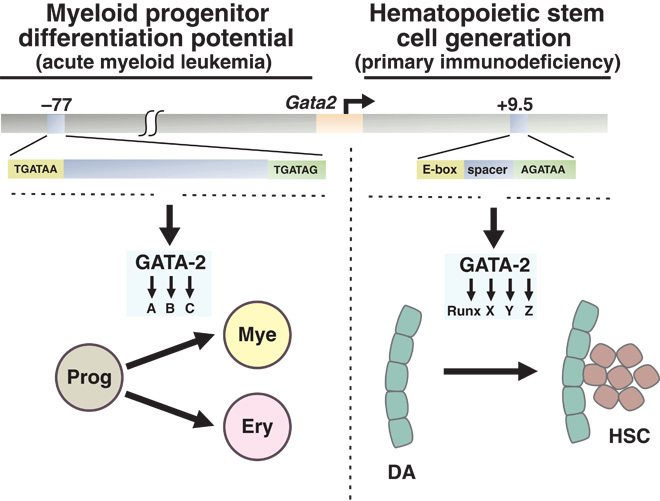
Cis-element encyclopedias provide information on phenotypic diversity and disease mechanisms. Although cis-element polymorphisms and mutations are instructive, deciphering function remains challenging. Mutation of an intronic GATA motif (+9.5) in GATA2, encoding a master regulator of hematopoiesis, underlies an immunodeficiency associated with myelodysplastic syndrome (MDS) and acute myeloid leukemia (AML). Whereas an inversion relocalizes another GATA2 cis-element (-77) to the proto-oncogene EVI1, inducing EVI1 expression and AML, whether this reflects ectopic or physiological activity is unknown. We describe a mouse strain that decouples -77 function from proto-oncogene deregulation. The -77(-/-) mice exhibited a novel phenotypic constellation including late embryonic lethality and anemia. The -77 established a vital sector of the myeloid progenitor transcriptome, conferring multipotentiality. Unlike the +9.5(-/-) embryos, hematopoietic stem cell genesis was unaffected in -77(-/-) embryos. These results illustrate a paradigm in which cis-elements in a locus differentially control stem and progenitor cell transitions, and therefore the individual cis-element alterations cause unique and overlapping disease phenotypes.
Keywords: GATA factor; GATA-2; cis-element; differentiation; hematopoiesis; myeloid; progenitor; stem cell; transcriptome.
Figures
References
-
- Yue F., Cheng Y., Breschi A., Vierstra J., Wu W., Ryba T., Sandstrom R., Ma Z., Davis C., Pope B. D., Shen Y., Pervouchine D. D., Djebali S., Thurman R. E., Kaul R., Rynes E., Kirilusha A., Marinov G. K., Williams B. A., Trout D., Amrhein H., Fisher-Aylor K., Antoshechkin I., DeSalvo G., See L.-H., Fastuca M., Drenkow J., Zaleski C., Dobin A., Prieto P., Lagarde J., Bussotti G., Tanzer A., Denas O., Li K., Bender M. A., Zhang M., Byron R., Groudine M. T., McCleary D., Pham L., Ye Z., Kuan S., Edsall L., Wu Y.-C., Rasmussen M. D., Bansal M. S., Kellis M., Keller C. A., Morrissey C. S., Mishra T., Jain D., Dogan N., Harris R. S., Cayting P., Kawli T., Boyle A. P., Euskirchen G., Kundaje A., Lin S., Lin Y., Jansen C., Malladi V. S., Cline M. S., Erickson D. T., Kirkup V. M., Learned K., Sloan C. A., Rosenbloom K. R., Lacerda de Sousa B., Beal K., Pignatelli M., Flicek P., Lian J., Kahveci T., Lee D., Kent W. J., Santos M. R., Herrero J., Notredame C., Johnson A., Vong S., Lee K., Bates D., Neri F., Diegel M., Canfield T., Sabo P. J., Wilken M. S., Reh T. A., Giste E., Shafer A., Kutyavin T., Haugen E., Dunn D., Reynolds A. P., Neph S., Humbert R., Scott Hansen R., De Bruijn M., Selleri L., Rudensky A., Josefowicz S., Samstein R., Eichler E. E., Orkin S. H., Levasseur D., Papayannopoulou T., Chang K.-H., Skoultchi A., Gosh S., Disteche C., Treuting P., Wang Y., Weiss M. J., Blobel G. A., Cao X., Zhong S., Wang T., Good P. J., Lowdon R. F., Adams L. B., Zhou X.-Q., Pazin M. J., Feingold E. A., Wold B., Taylor J., Mortazavi A., Weissman S. M., Stamatoyannopoulos J. A., Snyder M. P., Guigo R., Gingeras T. R., Gilbert D. M., Hardison R. C., Beer M. A., Ren, B. The Mouse ENCODE Consortium , A comparative encyclopedia of DNA elements in the mouse genome. Nature 515, 355–364 (2014). - PMC - PubMed
-
- Kellis M., Wold B., Snyder M. P., Bernstein B. E., Kundaje A., Marinov G. K., Ward L. D., Birney E., Crawford G. E., Dekker J., Dunham I., Elnitski L. L., Farnham P. J., Feingold E. A., Gerstein M., Giddings M. C., Gilbert D. M., Gingeras T. R., Green E. D., Guigo R., Hubbard T., Kent J., Lieb J. D., Myers R. M., Pazin M. J., Ren B., Stamatoyannopoulos J. A., Weng Z., White K. P., Hardison R. C., Defining functional DNA elements in the human genome. Proc. Natl. Acad. Sci. U.S.A. 111, 6131–6138 (2014). - PMC - PubMed
-
- Tsai F. Y., Keller G., Kuo F. C., Weiss M., Chen J., Rosenblatt M., Alt F. W., Orkin S. H., An early haematopoietic defect in mice lacking the transcription factor GATA-2. Nature 371, 221–226 (1994). - PubMed
-
- Tsai F.-Y., Orkin S. H., Transcription factor GATA-2 is required for proliferation/survival of early hematopoietic cells and mast cell formation, but not for erythroid and myeloid terminal differentiation. Blood 89, 3636–3643 (1997). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
