

Uncertainty in the Timing of Origin of Animals and the Limits of Precision in Molecular Timescales
- PMID: 26603774
- PMCID: PMC4651906
- DOI: 10.1016/j.cub.2015.09.066
Uncertainty in the Timing of Origin of Animals and the Limits of Precision in Molecular Timescales
Abstract
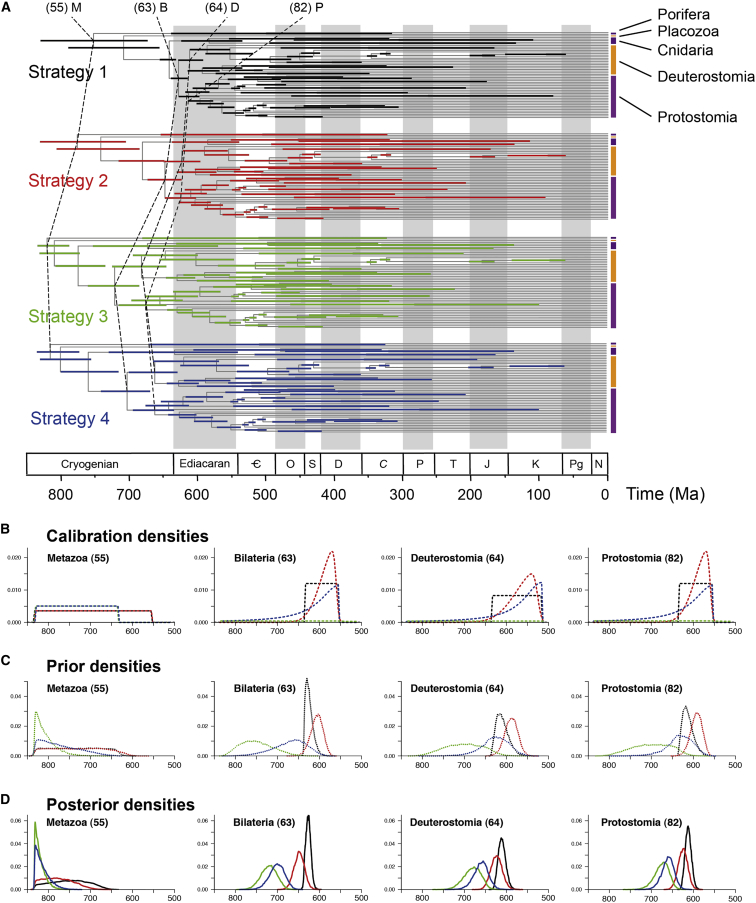
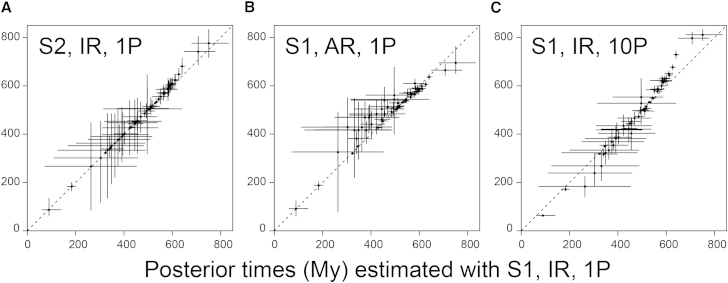
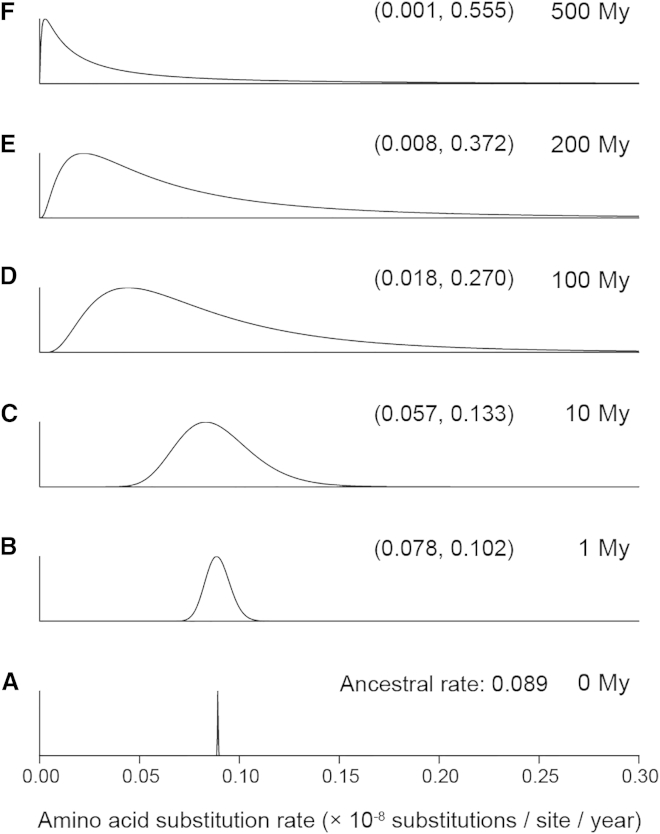
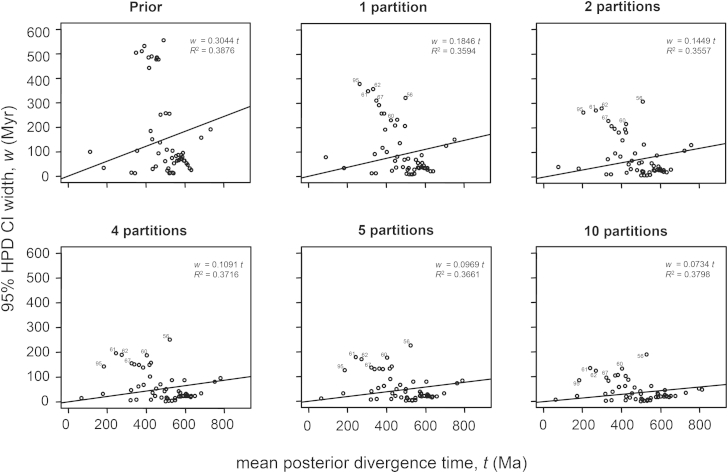
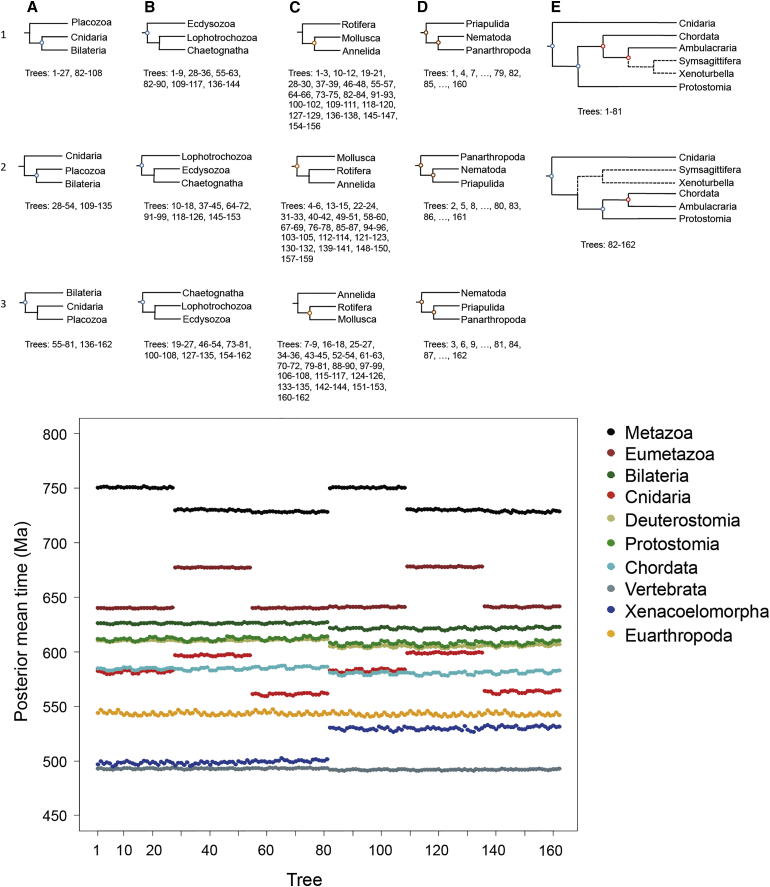
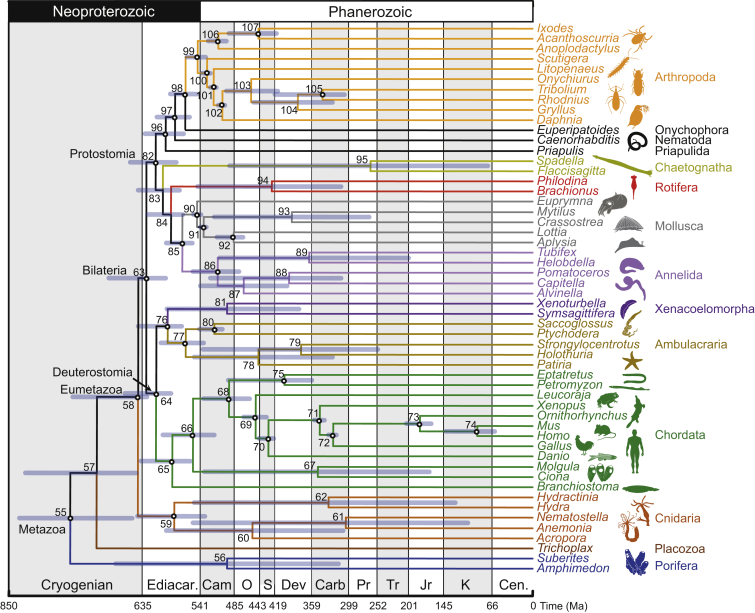
The timing of divergences among metazoan lineages is integral to understanding the processes of animal evolution, placing the biological events of species divergences into the correct geological timeframe. Recent fossil discoveries and molecular clock dating studies have suggested a divergence of bilaterian phyla >100 million years before the Cambrian, when the first definite crown-bilaterian fossils occur. Most previous molecular clock dating studies, however, have suffered from limited data and biases in methodologies, and virtually all have failed to acknowledge the large uncertainties associated with the fossil record of early animals, leading to inconsistent estimates among studies. Here we use an unprecedented amount of molecular data, combined with four fossil calibration strategies (reflecting disparate and controversial interpretations of the metazoan fossil record) to obtain Bayesian estimates of metazoan divergence times. Our results indicate that the uncertain nature of ancient fossils and violations of the molecular clock impose a limit on the precision that can be achieved in estimates of ancient molecular timescales. For example, although we can assert that crown Metazoa originated during the Cryogenian (with most crown-bilaterian phyla diversifying during the Ediacaran), it is not possible with current data to pinpoint the divergence events with sufficient accuracy to test for correlations between geological and biological events in the history of animals. Although a Cryogenian origin of crown Metazoa agrees with current geological interpretations, the divergence dates of the bilaterians remain controversial. Thus, attempts to build evolutionary narratives of early animal evolution based on molecular clock timescales appear to be premature.
Figures
Comment in
-
Animal Evolution: Only Rocks Can Set the Clock.Curr Biol. 2015 Nov 16;25(22):R1079-81. doi: 10.1016/j.cub.2015.10.015. Curr Biol. 2015. PMID: 26583898
References
-
- Signor P.W., Lipps J.H. Origin and early radiation of the Metazoa. In: Signor P.W., Lipps J.H., editors. Origin and Early Evolution of the Metazoa. Springer; 1992. pp. 3–23.
-
- Planavsky N.J., Reinhard C.T., Wang X., Thomson D., McGoldrick P., Rainbird R.H., Johnson T., Fischer W.W., Lyons T.W. Earth history. Low mid-Proterozoic atmospheric oxygen levels and the delayed rise of animals. Science. 2014;346:635–638. - PubMed
-
- Knoll A.H., Carroll S.B. Early animal evolution: emerging views from comparative biology and geology. Science. 1999;284:2129–2137. - PubMed
-
- Hoffman P.F., Schrag D.P. The snowball Earth hypothesis: testing the limits of global change. Terra Nova. 2002;14:129–155.
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
