

Common features in diverse insect clocks
- PMID: 26605055
- PMCID: PMC4604113
- DOI: 10.1186/s40851-014-0003-y
Common features in diverse insect clocks
Abstract
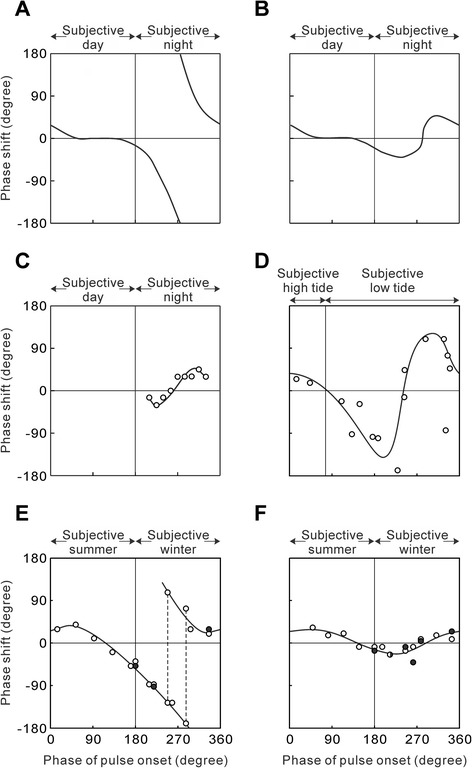
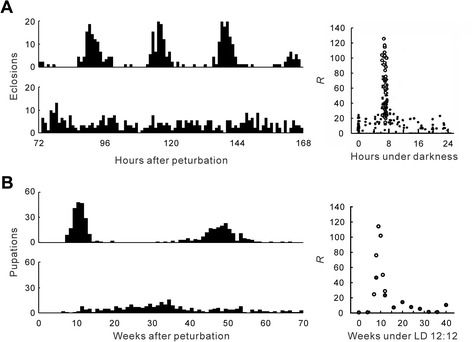
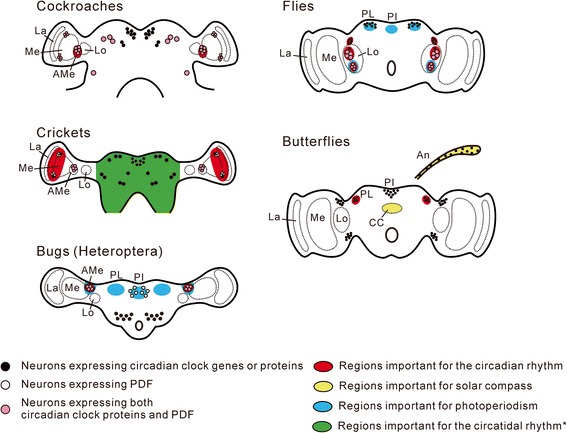
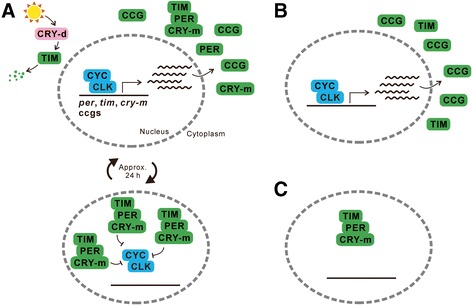
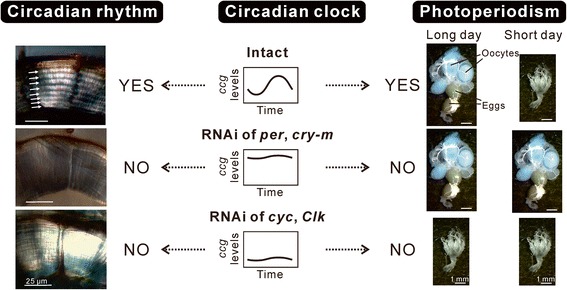
This review describes common features among diverse biological clocks in insects, including circadian, circatidal, circalunar/circasemilunar, and circannual clocks. These clocks control various behaviors, physiological functions, and developmental events, enabling adaptation to periodic environmental changes. Circadian clocks also function in time-compensation for celestial navigation and in the measurement of day or night length for photoperiodism. Phase response curves for such clocks reported thus far exhibit close similarities; specifically, the circannual clock in Anthrenus verbasci shows striking similarity to circadian clocks in its phase response. It is suggested that diverse biological clocks share physiological properties in their phase responses irrespective of period length. Molecular and physiological mechanisms are best understood for the optic-lobe and mid-brain circadian clocks, although there is no direct evidence that these clocks are involved in rhythmic phenomena other than circadian rhythms in daily events. Circadian clocks have also been localized in peripheral tissues, and research on their role in various rhythmic phenomena has been started. Although clock genes have been identified as controllers of circadian rhythms in daily events, some of these genes have also been shown to be involved in photoperiodism and possibly in time-compensated celestial navigation. In contrast, there is no experimental evidence indicating that any known clock gene is involved in biological clocks other than circadian clocks.
Keywords: Anatomical location; Celestial navigation; Circadian; Circalunar; Circannual; Circasemilunar; Circatidal; Clock gene; Phase response curve; Photoperiodism.
Figures
References
-
- Brown FA, Jr, Hastings JW, Palmer JD. The Biological Clock - Two Views. New York: Academic Press; 1970.
-
- Saunders DS. Insect Clocks. 3. Amsterdam: Elsevier; 2002.
-
- Pittendrigh CS. The circadian oscillation in Drosophila pseudoobscura pupae: a model for the photoperiodic clock. Z Pflanzenphysiol. 1966;54:275–307.
-
- von Frisch K. Gelöste und ungelöste Rätsel der Bienensprache. Naturwissenschaften. 1948;35:12–23. doi: 10.1007/BF00626624. - DOI
-
- Perez SM, Taylor OR, Jander R. A sun compass in monarch butterflies. Nature. 1997;387:29. doi: 10.1038/387029a0. - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
