

Rescue from tau-induced neuronal dysfunction produces insoluble tau oligomers
- PMID: 26608845
- PMCID: PMC4660438
- DOI: 10.1038/srep17191
Rescue from tau-induced neuronal dysfunction produces insoluble tau oligomers
Abstract
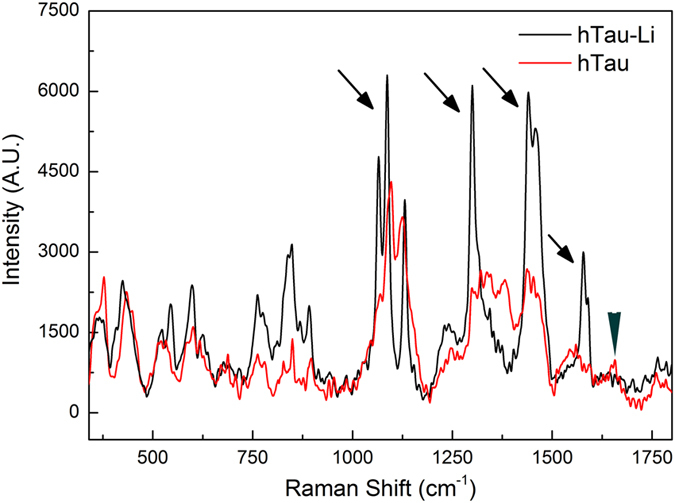
Aggregation of highly phosphorylated tau is a hallmark of Alzheimer's disease and other tauopathies. Nevertheless, animal models demonstrate that tau-mediated dysfunction/toxicity may not require large tau aggregates but instead may be caused by soluble hyper-phosphorylated tau or by small tau oligomers. Challenging this widely held view, we use multiple techniques to show that insoluble tau oligomers form in conditions where tau-mediated dysfunction is rescued in vivo. This shows that tau oligomers are not necessarily always toxic. Furthermore, their formation correlates with increased tau levels, caused intriguingly, by either pharmacological or genetic inhibition of tau kinase glycogen-synthase-kinase-3beta (GSK-3β). Moreover, contrary to common belief, these tau oligomers were neither highly phosphorylated, and nor did they contain beta-pleated sheet structure. This may explain their lack of toxicity. Our study makes the novel observation that tau also forms non-toxic insoluble oligomers in vivo in addition to toxic oligomers, which have been reported by others. Whether these are inert or actively protective remains to be established. Nevertheless, this has wide implications for emerging therapeutic strategies such as those that target dissolution of tau oligomers as they may be ineffective or even counterproductive unless they act on the relevant toxic oligomeric tau species.
Figures
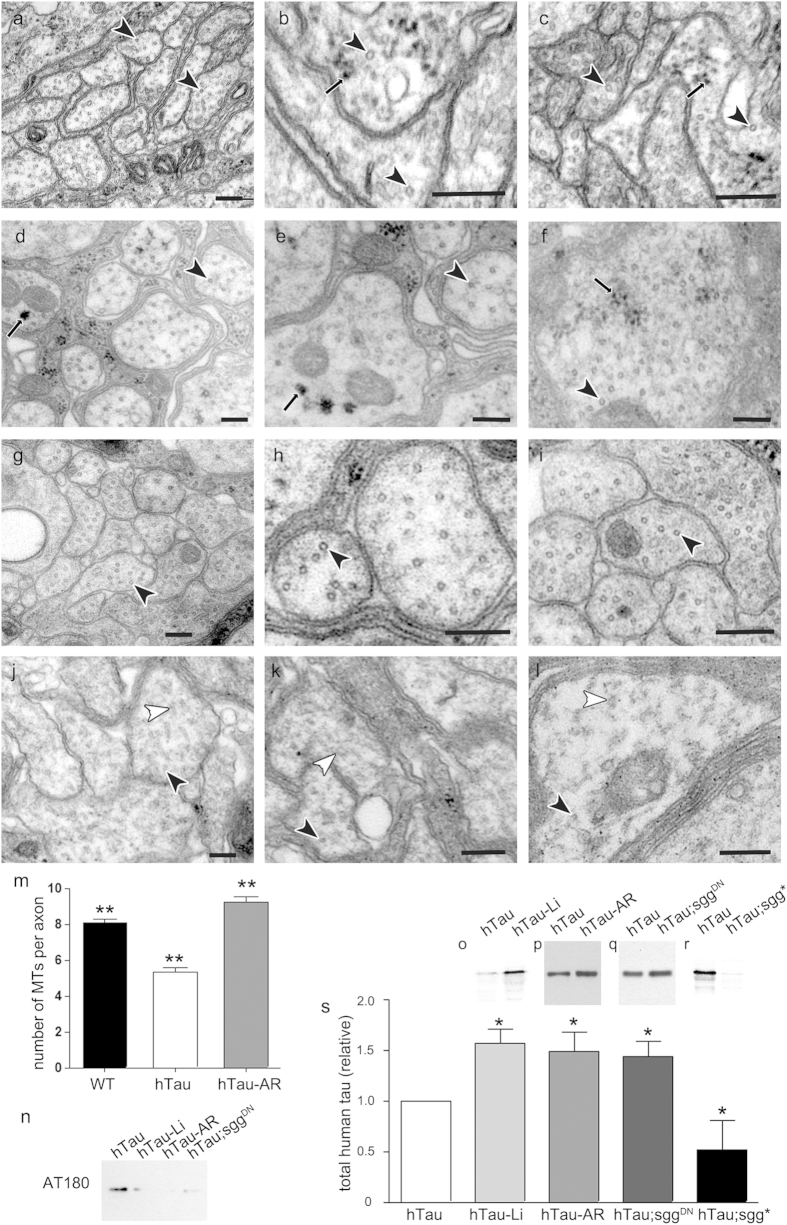
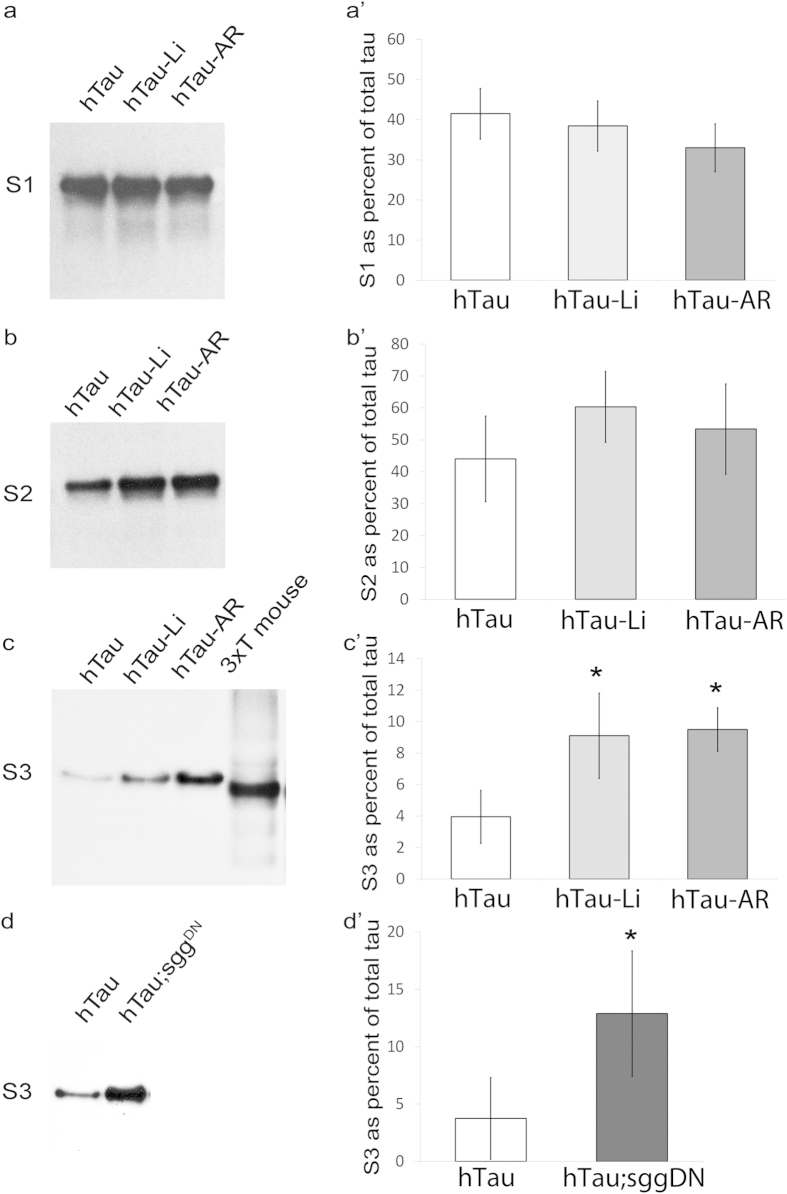
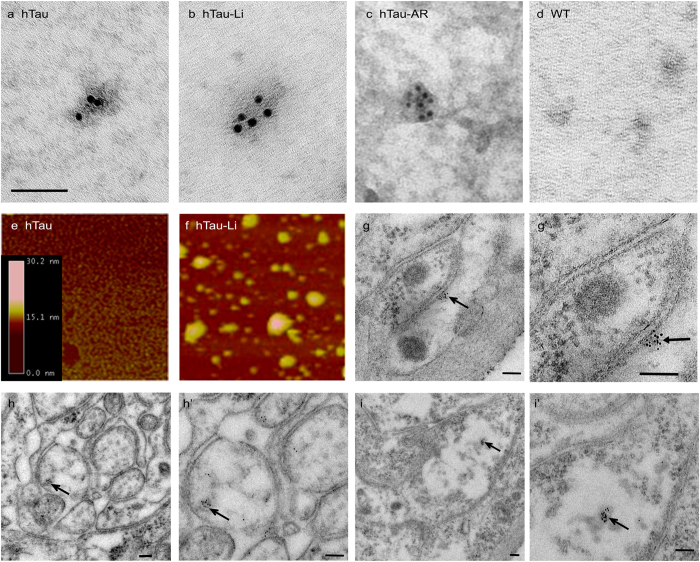
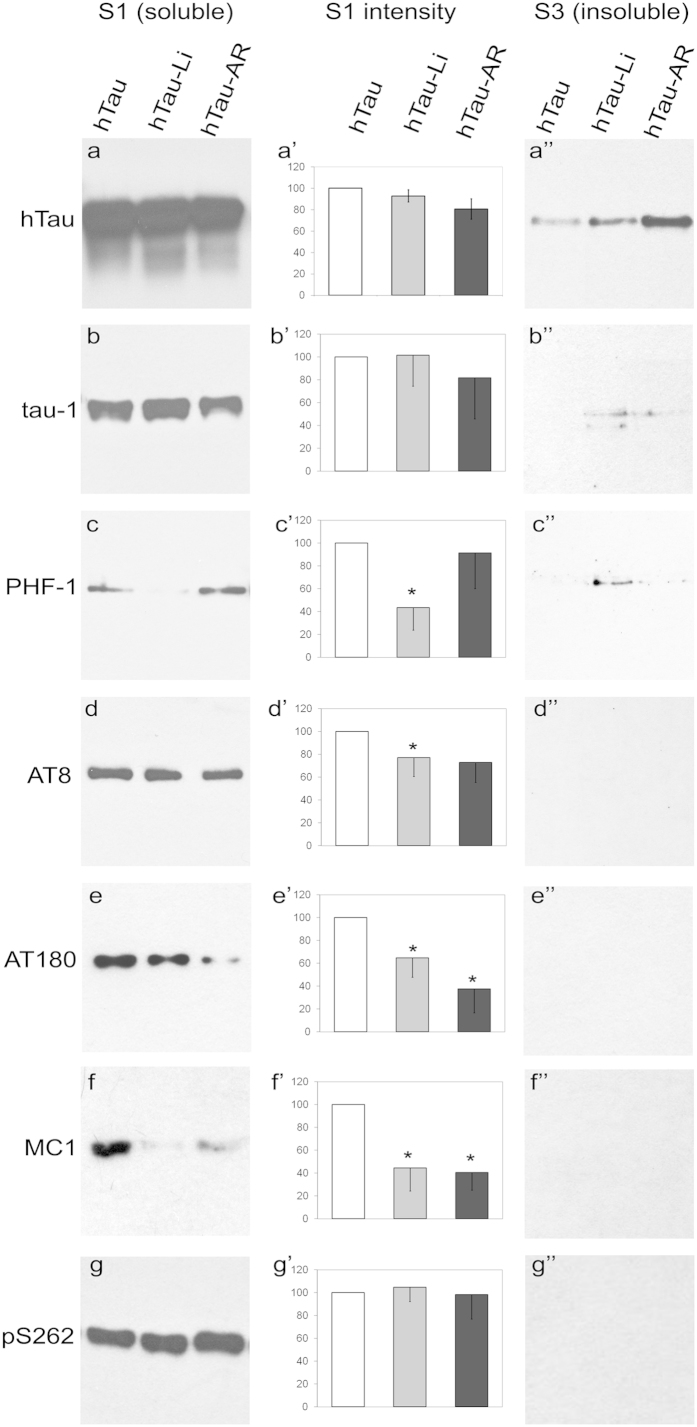
References
-
- Hanger D. P., Hughes K., Woodgett J. R., Brion J. P. & Anderton B. H. Glycogen synthase kinase-3 induces Alzheimer’s disease-like phosphorylation of tau: generation of paired helical filament epitopes and neuronal localisation of the kinase. Neurosci. Lett. 147, 58–62 (1992). - PubMed
-
- Lovestone S. et al. Alzheimer’s disease-like phosphorylation of the microtubule-associated protein tau by glycogen synthase kinase-3 in transfected mammalian cells. Curr. Biol. 4, 1077–1086 (1994). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
