

Intein Clustering Suggests Functional Importance in Different Domains of Life
- PMID: 26609079
- PMCID: PMC4760082
- DOI: 10.1093/molbev/msv271
Intein Clustering Suggests Functional Importance in Different Domains of Life
Abstract
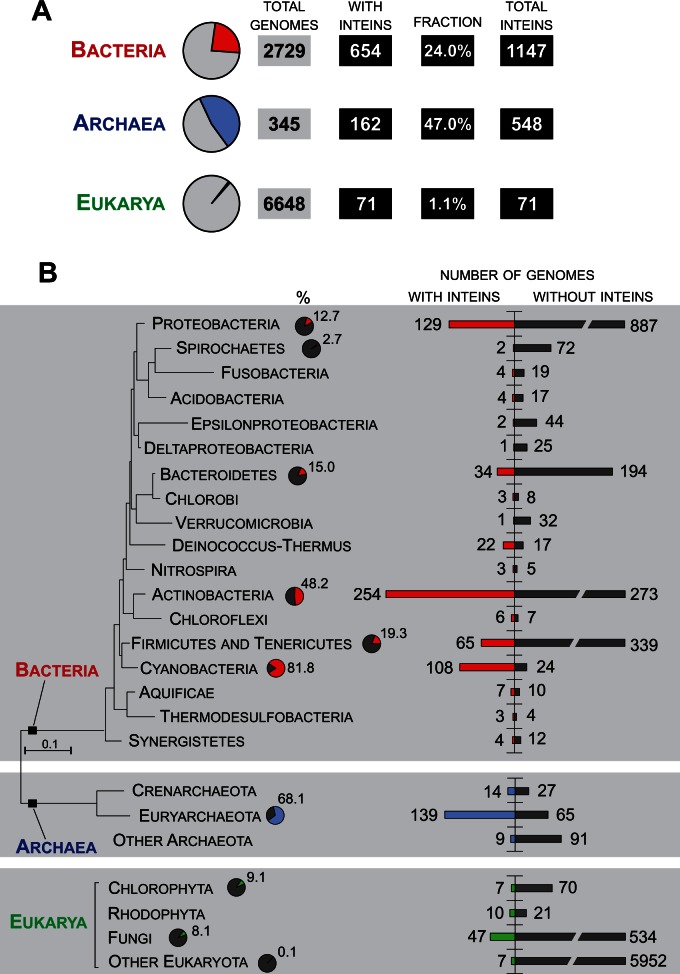
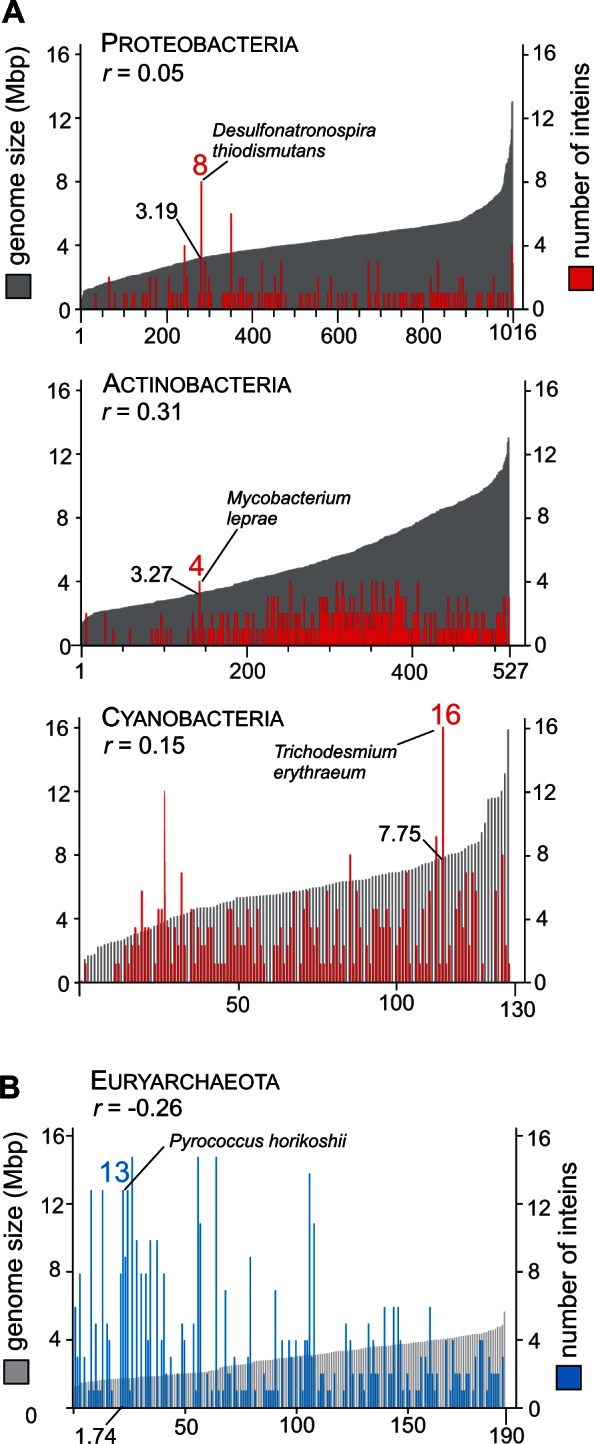
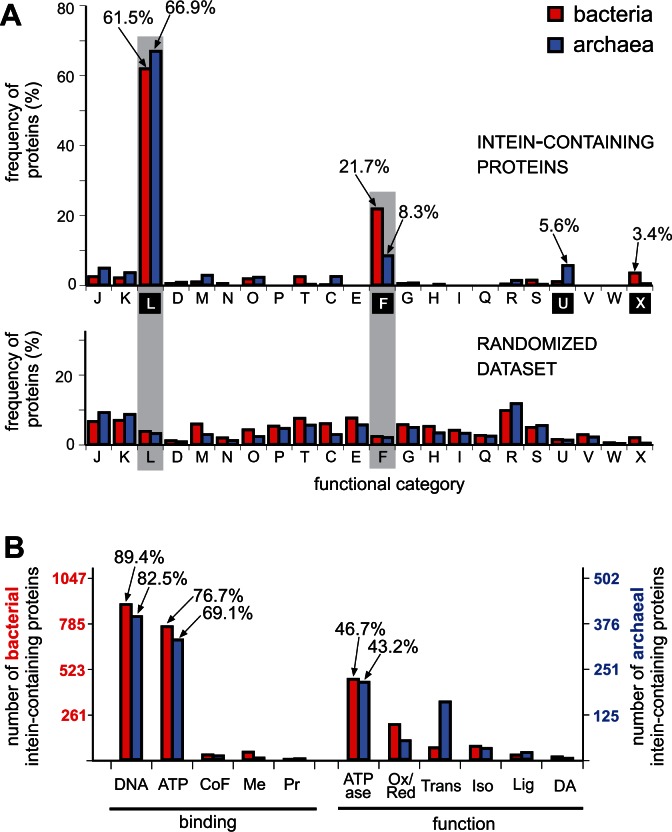
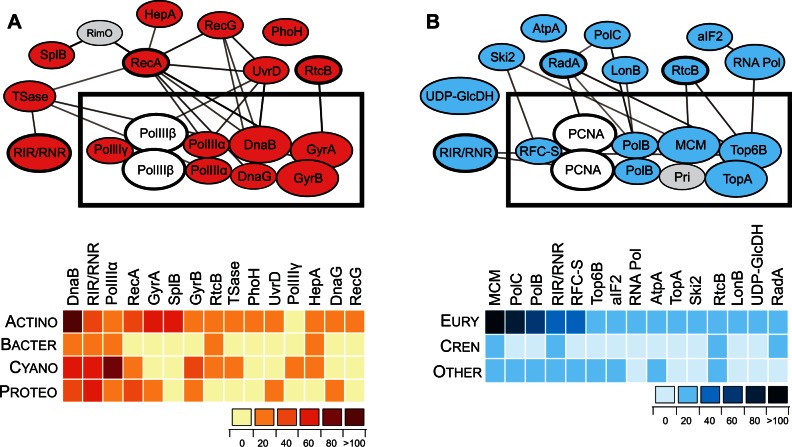
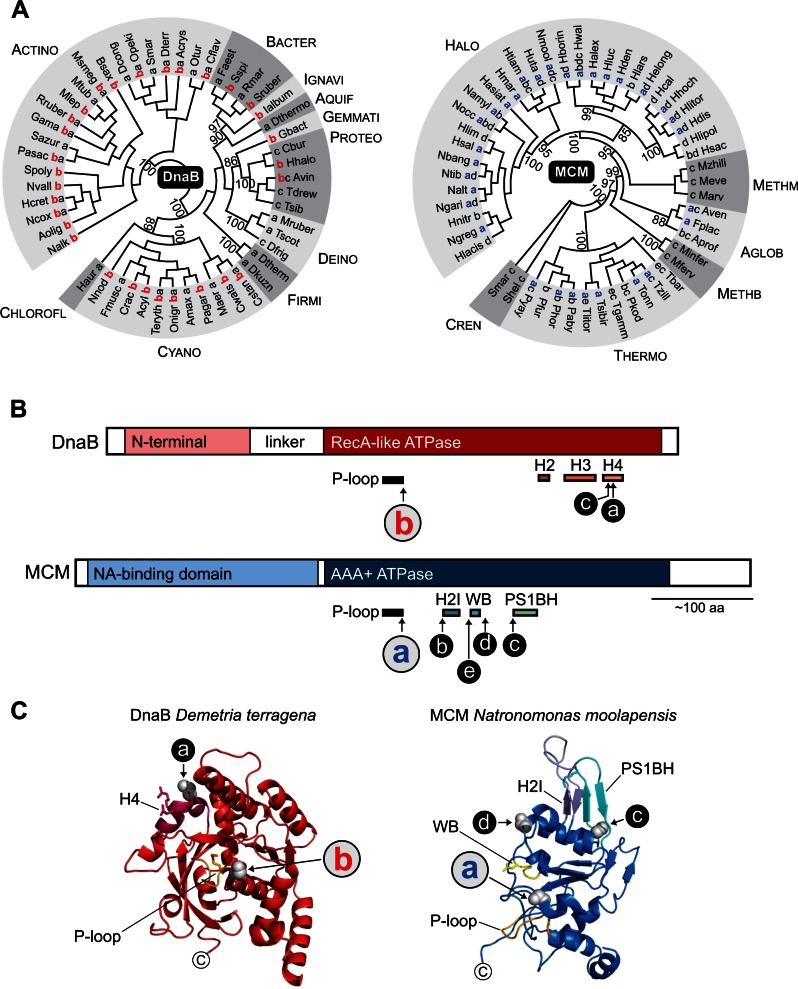
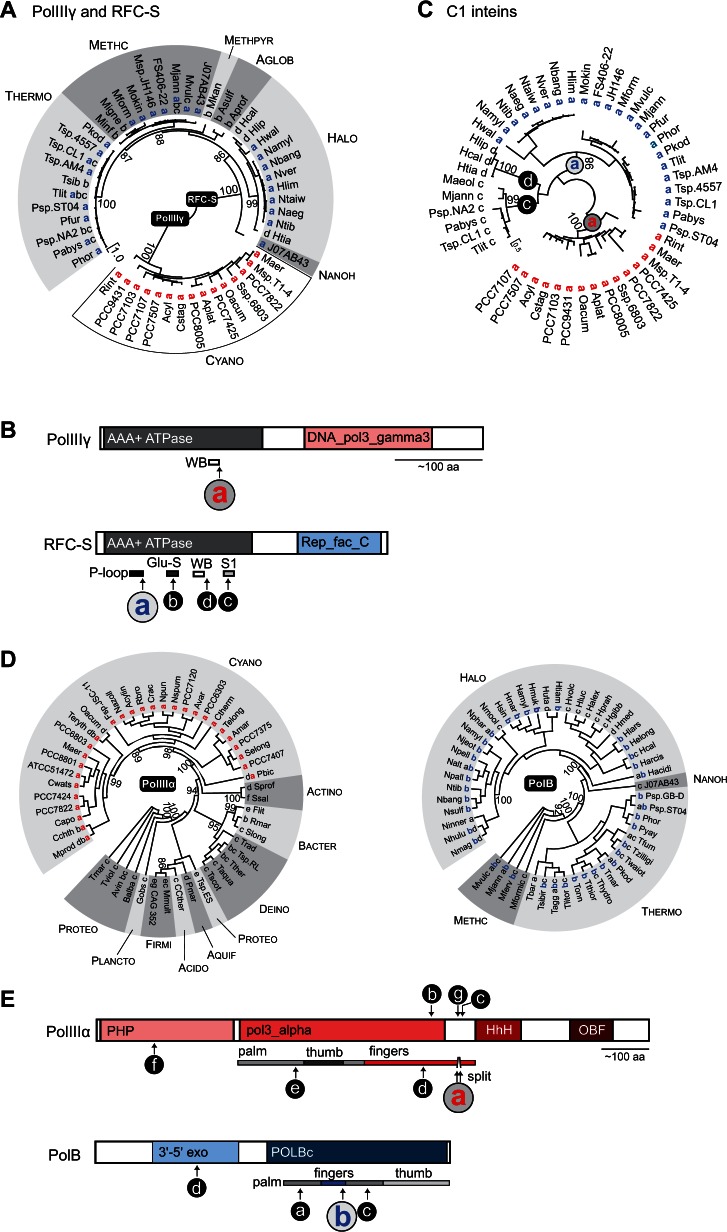
Inteins, also called protein introns, are self-splicing mobile elements found in all domains of life. A bioinformatic survey of genomic data highlights a biased distribution of inteins among functional categories of proteins in both bacteria and archaea, with a strong preference for a single network of functions containing replisome proteins. Many nonorthologous, functionally equivalent replicative proteins in bacteria and archaea carry inteins, suggesting a selective retention of inteins in proteins of particular functions across domains of life. Inteins cluster not only in proteins with related roles but also in specific functional units of those proteins, like ATPase domains. This peculiar bias does not fully fit the models describing inteins exclusively as parasitic elements. In such models, evolutionary dynamics of inteins is viewed primarily through their mobility with the intein homing endonuclease (HEN) as the major factor of intein acquisition and loss. Although the HEN is essential for intein invasion and spread in populations, HEN dynamics does not explain the observed biased distribution of inteins among proteins in specific functional categories. We propose that the protein splicing domain of the intein can act as an environmental sensor that adapts to a particular niche and could increase the chance of the intein becoming fixed in a population. We argue that selective retention of some inteins might be beneficial under certain environmental stresses, to act as panic buttons that reversibly inhibit specific networks, consistent with the observed intein distribution.
Keywords: ATPases; Clusters of Orthologous Groups; evolution; replicative helicase; replisome.
© The Author(s) 2015. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Allen MJ, Lanzén A, Bratbak G. 2011. Characterisation of the coccolithovirus intein. Mar Genomics. 4:1–7. - PubMed
-
- Amitai G, Dassa B, Pietrokovski S. 2004. Protein splicing of inteins with atypical glutamine and aspartate C-terminal residues. J Biol Chem. 279:3121–3131. - PubMed
-
- Anisimova M, Gascuel O. 2006. Approximate likelihood-ratio test for branches: a fast, accurate, and powerful alternative. Syst Biol. 55:539–552. - PubMed
-
- Barzel A, Naor A, Privman E, Kupiec M, Gophna U. 2011. Homing endonucleases residing within inteins: evolutionary puzzles awaiting genetic solutions. Biochem Soc Trans. 39:169–173. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
