

Active Dendrites and Differential Distribution of Calcium Channels Enable Functional Compartmentalization of Golgi Cells
- PMID: 26609148
- PMCID: PMC4659820
- DOI: 10.1523/JNEUROSCI.3132-15.2015
Active Dendrites and Differential Distribution of Calcium Channels Enable Functional Compartmentalization of Golgi Cells
Abstract
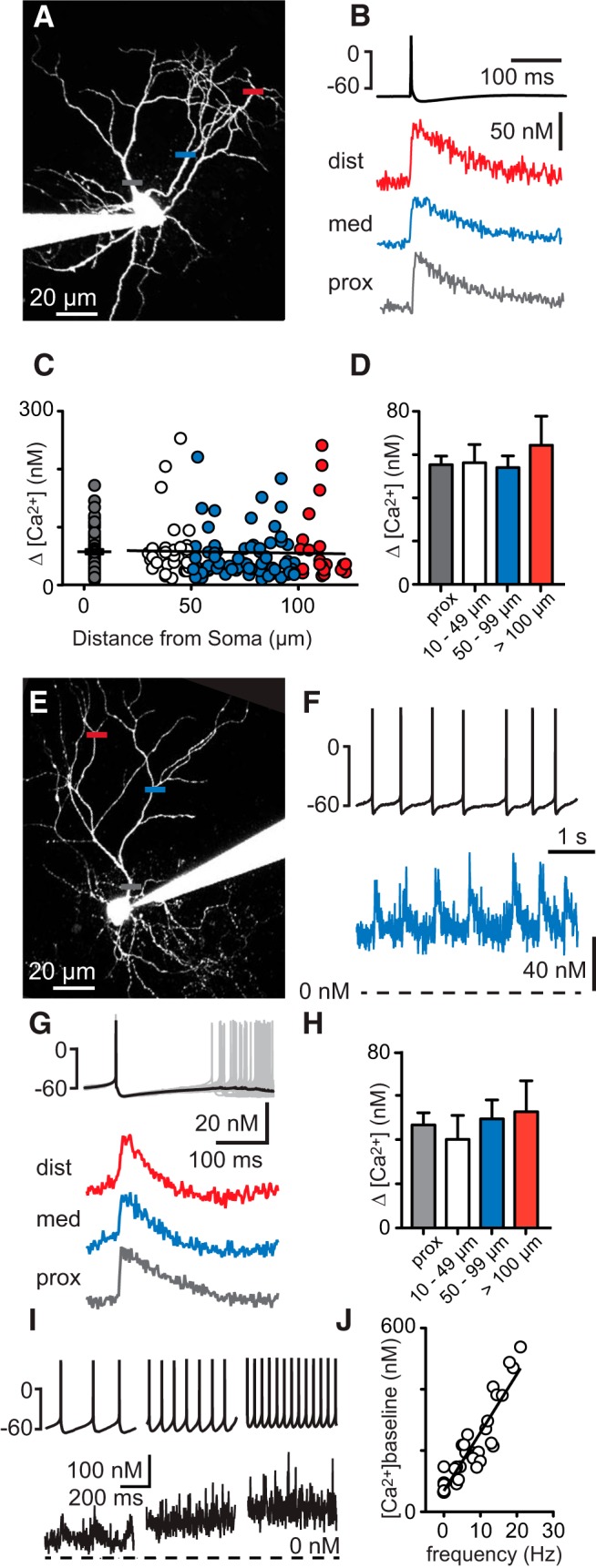
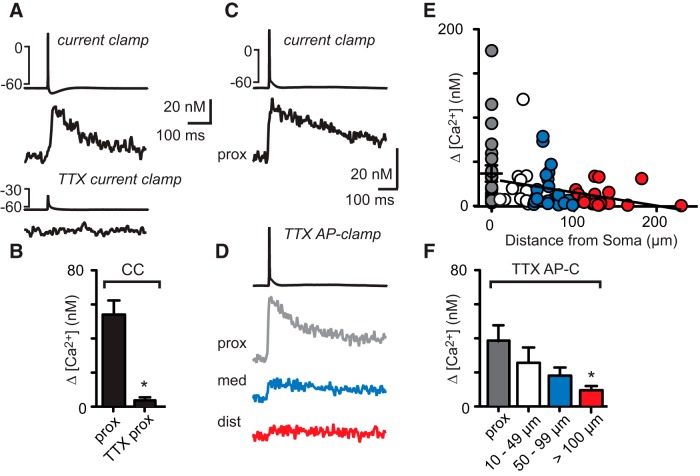
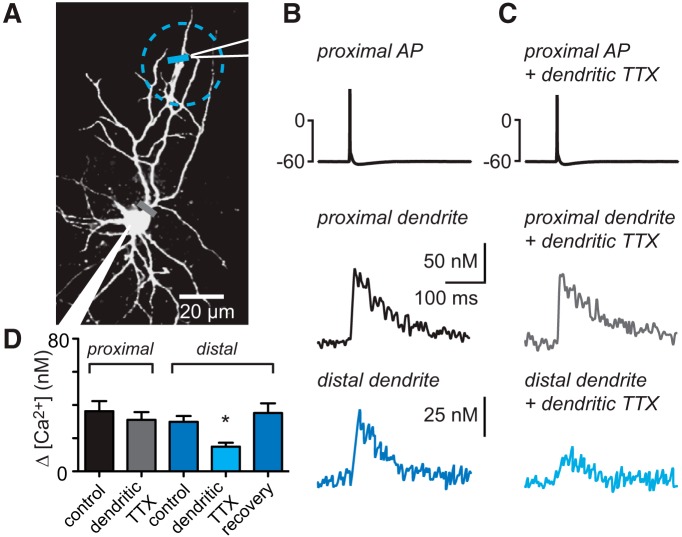
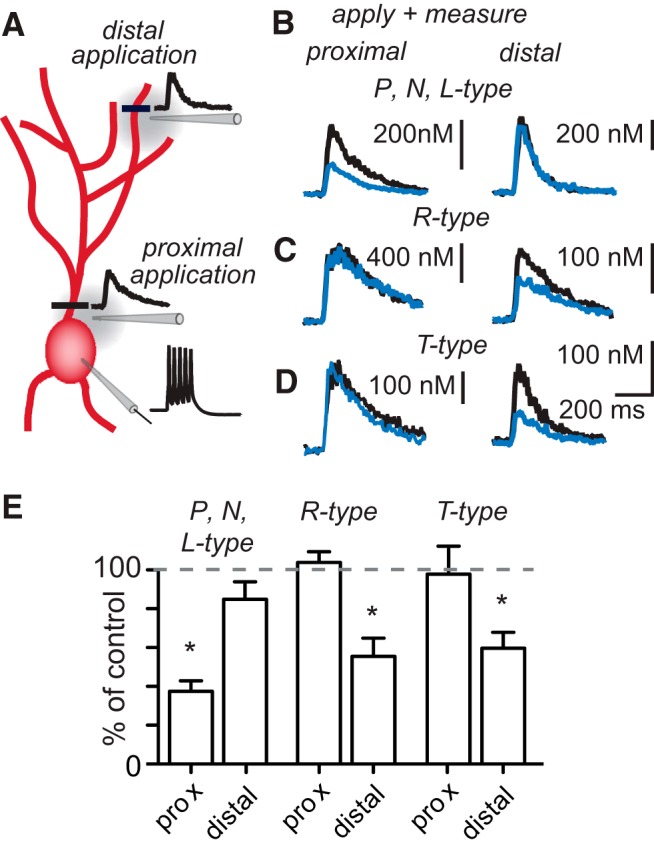
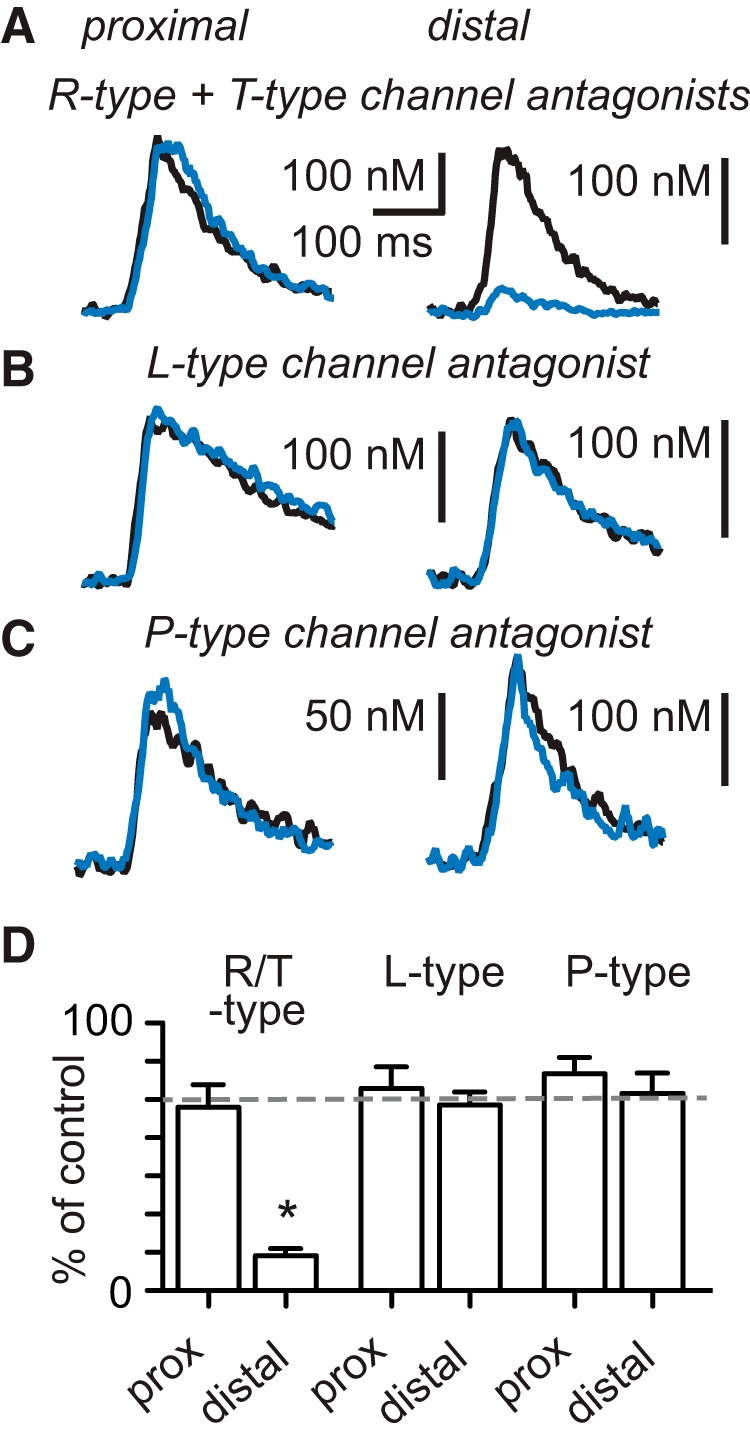
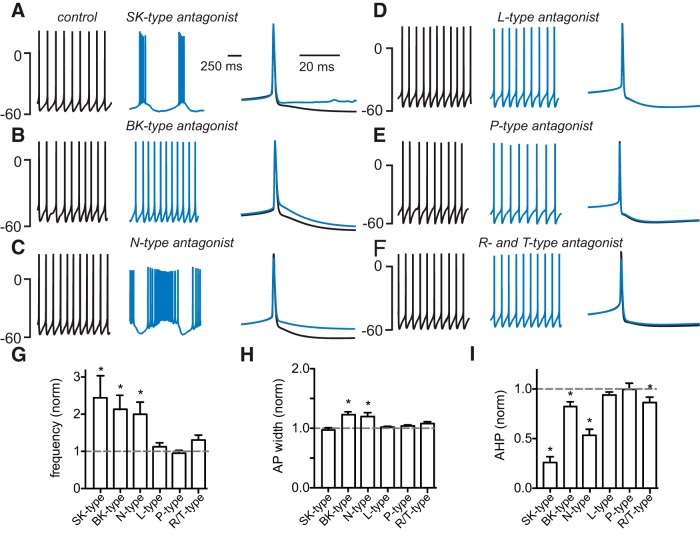
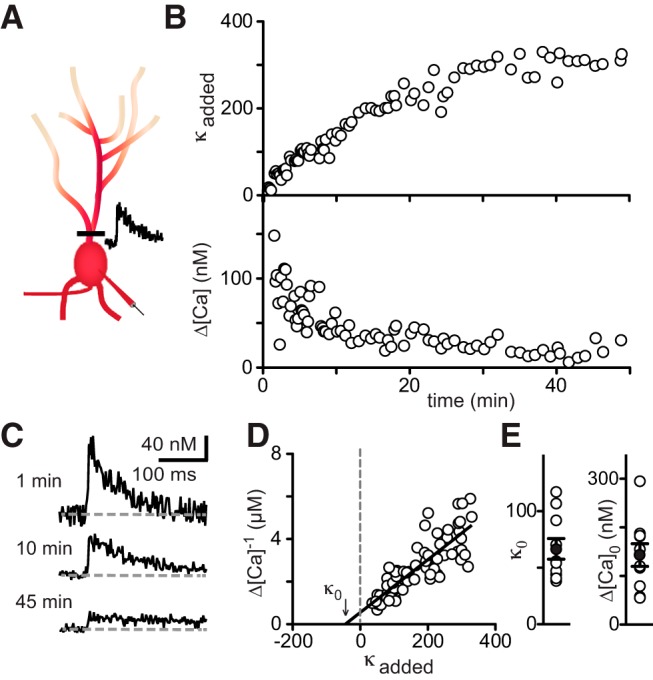
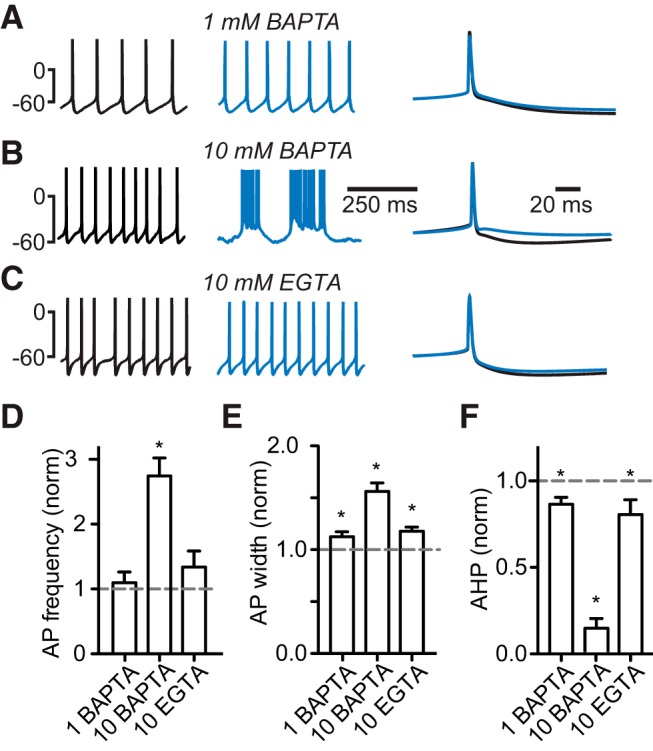
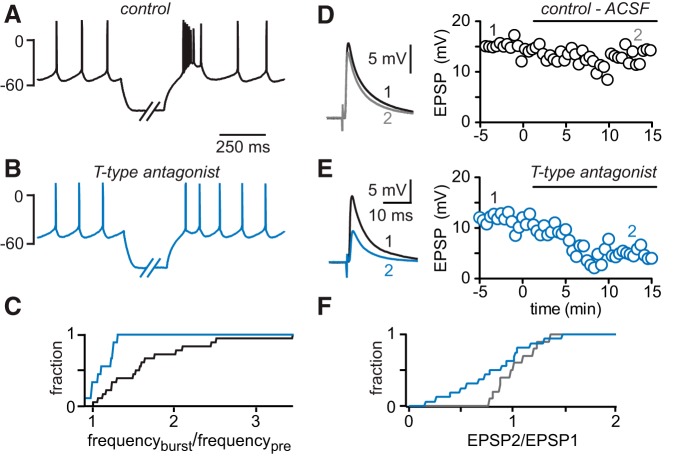
Interneurons are essential to controlling excitability, timing, and synaptic integration in neuronal networks. Golgi cells (GoCs) serve these roles at the input layer of the cerebellar cortex by releasing GABA to inhibit granule cells (grcs). GoCs are excited by mossy fibers (MFs) and grcs and provide feedforward and feedback inhibition to grcs. Here we investigate two important aspects of GoC physiology: the properties of GoC dendrites and the role of calcium signaling in regulating GoC spontaneous activity. Although GoC dendrites are extensive, previous studies concluded they are devoid of voltage-gated ion channels. Hence, the current view holds that somatic voltage signals decay passively within GoC dendrites, and grc synapses onto distal dendrites are not amplified and are therefore ineffective at firing GoCs because of strong passive attenuation. Using whole-cell recording and calcium imaging in rat slices, we find that dendritic voltage-gated sodium channels allow somatic action potentials to activate voltage-gated calcium channels (VGCCs) along the entire dendritic length, with R-type and T-type VGCCs preferentially located distally. We show that R- and T-type VGCCs located in the dendrites can boost distal synaptic inputs and promote burst firing. Active dendrites are thus critical to the regulation of GoC activity, and consequently, to the processing of input to the cerebellar cortex. In contrast, we find that N-type channels are preferentially located near the soma, and control the frequency and pattern of spontaneous firing through their close association with calcium-activated potassium (KCa) channels. Thus, VGCC types are differentially distributed and serve specialized functions within GoCs.
Significance statement: Interneurons are essential to neural processing because they modulate excitability, timing, and synaptic integration within circuits. At the input layer of the cerebellar cortex, a single type of interneuron, the Golgi cell (GoC), carries these functions. The extent of inhibition depends on both spontaneous activity of GoCs and the excitatory synaptic input they receive. In this study, we find that different types of calcium channels are differentially distributed, with dendritic calcium channels being activated by somatic activity, boosting synaptic inputs and enabling bursting, and somatic calcium cannels promoting regular firing. We therefore challenge the current view that GoC dendrites are passive and identify the mechanisms that contribute to GoCs regulating the flow of sensory information in the cerebellar cortex.
Keywords: calcium buffering; calcium channels; calcium-activated potassium channels; cerebellum; dendritic excitability; interneuron.
Copyright © 2015 the authors 0270-6474/15/3515492-13$15.00/0.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Pathway-Specific Drive of Cerebellar Golgi Cells Reveals Integrative Rules of Cortical Inhibition.J Neurosci. 2019 Feb 13;39(7):1169-1181. doi: 10.1523/JNEUROSCI.1448-18.2018. Epub 2018 Dec 26. J Neurosci. 2019. PMID: 30587539 Free PMC article.
-
Granule cell ascending axon excitatory synapses onto Golgi cells implement a potent feedback circuit in the cerebellar granular layer.J Neurosci. 2013 Jul 24;33(30):12430-46. doi: 10.1523/JNEUROSCI.4897-11.2013. J Neurosci. 2013. PMID: 23884948 Free PMC article.
-
Dendritic voltage and calcium-gated channels amplify the variability of postsynaptic responses in a Purkinje cell model.J Neurophysiol. 1998 Aug;80(2):504-19. doi: 10.1152/jn.1998.80.2.504. J Neurophysiol. 1998. PMID: 9705446
-
How voltage-gated ion channels alter the functional properties of ganglion and amacrine cell dendrites.Arch Ital Biol. 2002 Oct;140(4):347-59. Arch Ital Biol. 2002. PMID: 12228988 Review.
-
Roles of ion channels in EPSP integration at neuronal dendrites.Neurosci Res. 2000 Jul;37(3):167-71. doi: 10.1016/s0168-0102(00)00120-6. Neurosci Res. 2000. PMID: 10940450 Review.
Cited by
-
Modeling the Cerebellar Microcircuit: New Strategies for a Long-Standing Issue.Front Cell Neurosci. 2016 Jul 8;10:176. doi: 10.3389/fncel.2016.00176. eCollection 2016. Front Cell Neurosci. 2016. PMID: 27458345 Free PMC article. Review.
-
Cerebellar Golgi cell models predict dendritic processing and mechanisms of synaptic plasticity.PLoS Comput Biol. 2020 Dec 30;16(12):e1007937. doi: 10.1371/journal.pcbi.1007937. eCollection 2020 Dec. PLoS Comput Biol. 2020. PMID: 33378395 Free PMC article.
-
Surface dynamics of voltage-gated ion channels.Channels (Austin). 2016 Jul 3;10(4):267-81. doi: 10.1080/19336950.2016.1153210. Epub 2016 Feb 18. Channels (Austin). 2016. PMID: 26891382 Free PMC article. Review.
-
Only a Minority of the Inhibitory Inputs to Cerebellar Golgi Cells Originates from Local GABAergic Cells.eNeuro. 2016 May 23;3(2):ENEURO.0055-16.2016. doi: 10.1523/ENEURO.0055-16.2016. eCollection 2016 Mar-Apr. eNeuro. 2016. PMID: 27257627 Free PMC article.
-
Pathway-Specific Drive of Cerebellar Golgi Cells Reveals Integrative Rules of Cortical Inhibition.J Neurosci. 2019 Feb 13;39(7):1169-1181. doi: 10.1523/JNEUROSCI.1448-18.2018. Epub 2018 Dec 26. J Neurosci. 2019. PMID: 30587539 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources