

SAGA DUB-Ubp8 Deubiquitylates Centromeric Histone Variant Cse4
- PMID: 26613948
- PMCID: PMC4751549
- DOI: 10.1534/g3.115.024877
SAGA DUB-Ubp8 Deubiquitylates Centromeric Histone Variant Cse4
Abstract
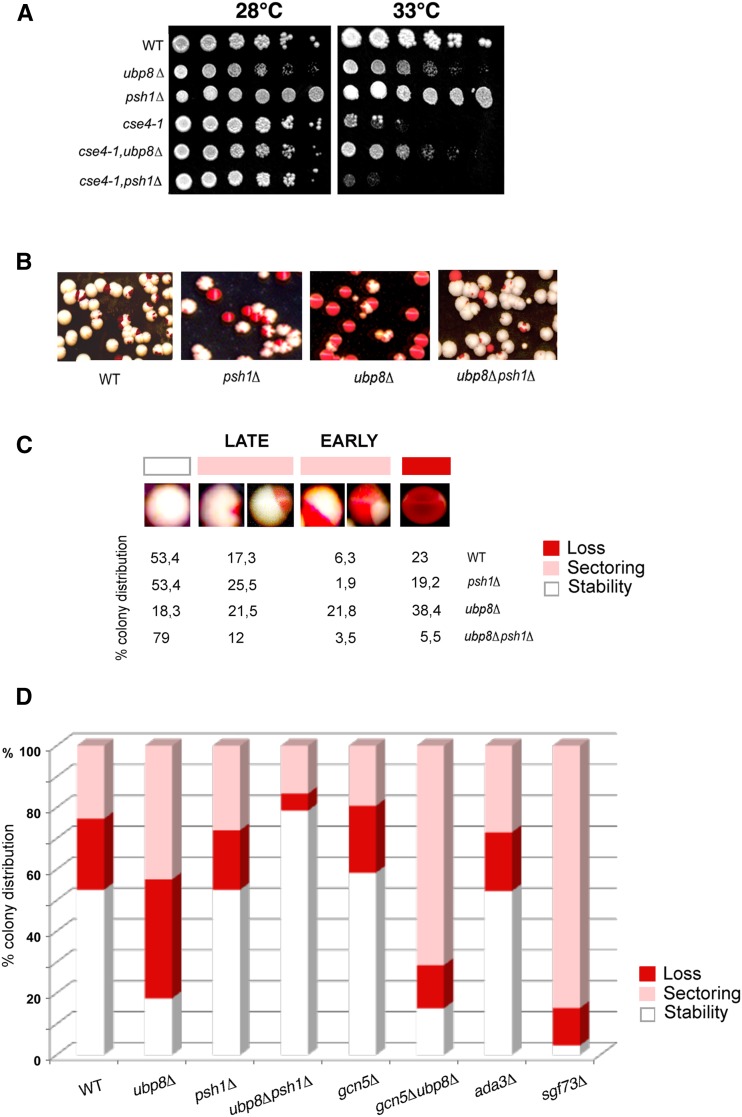
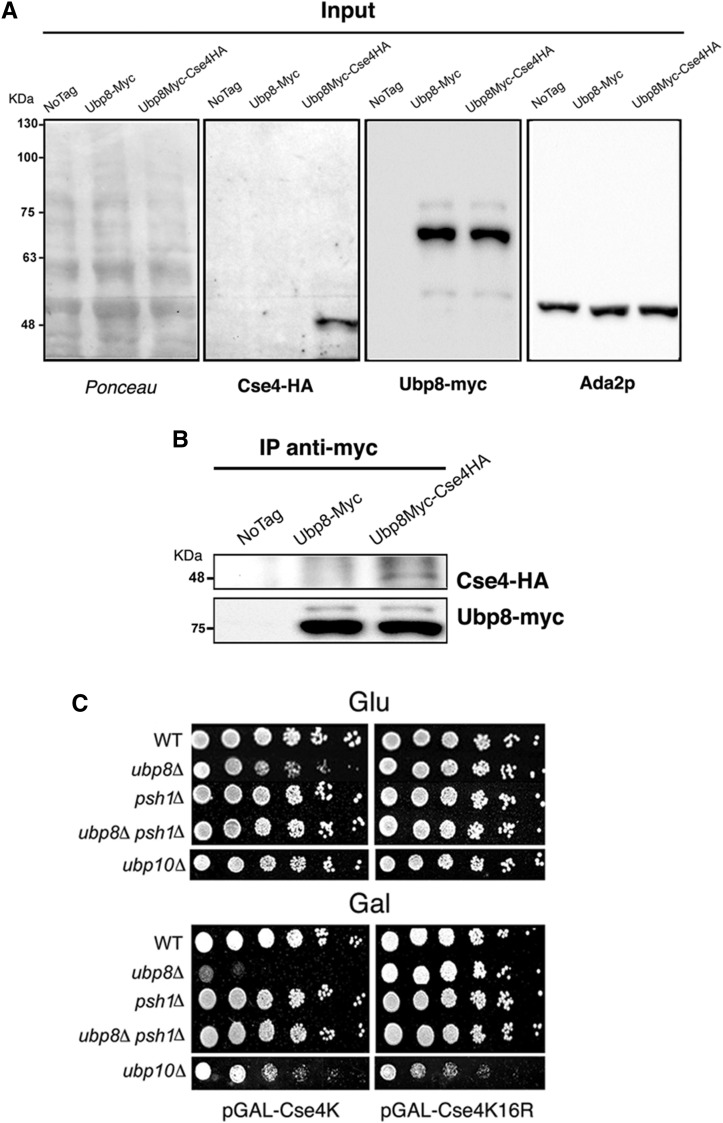
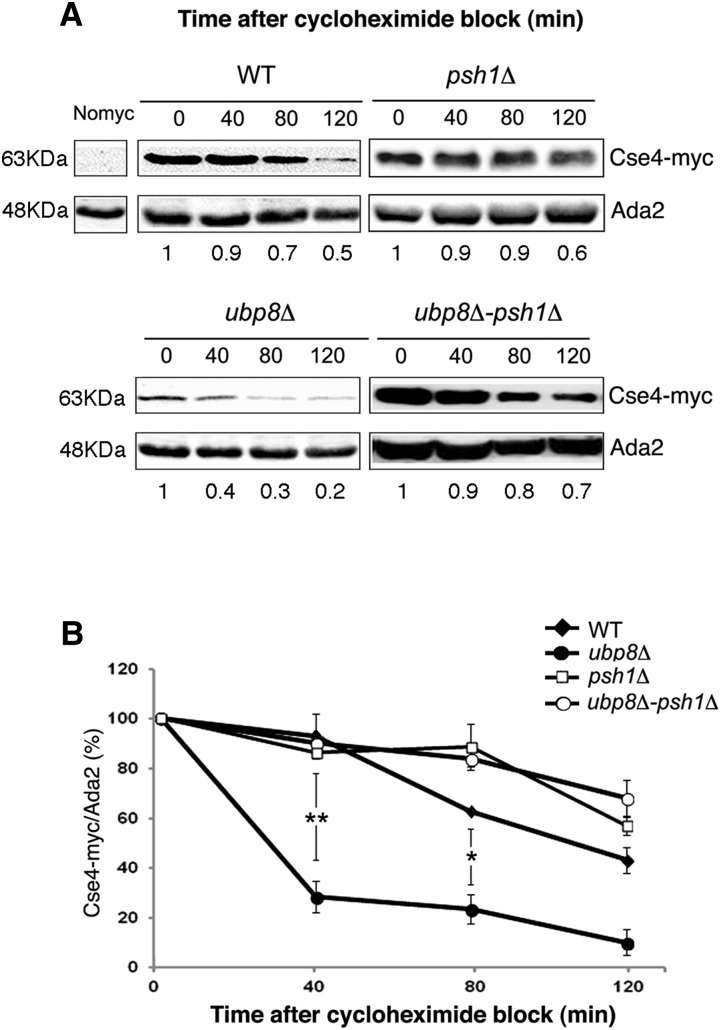
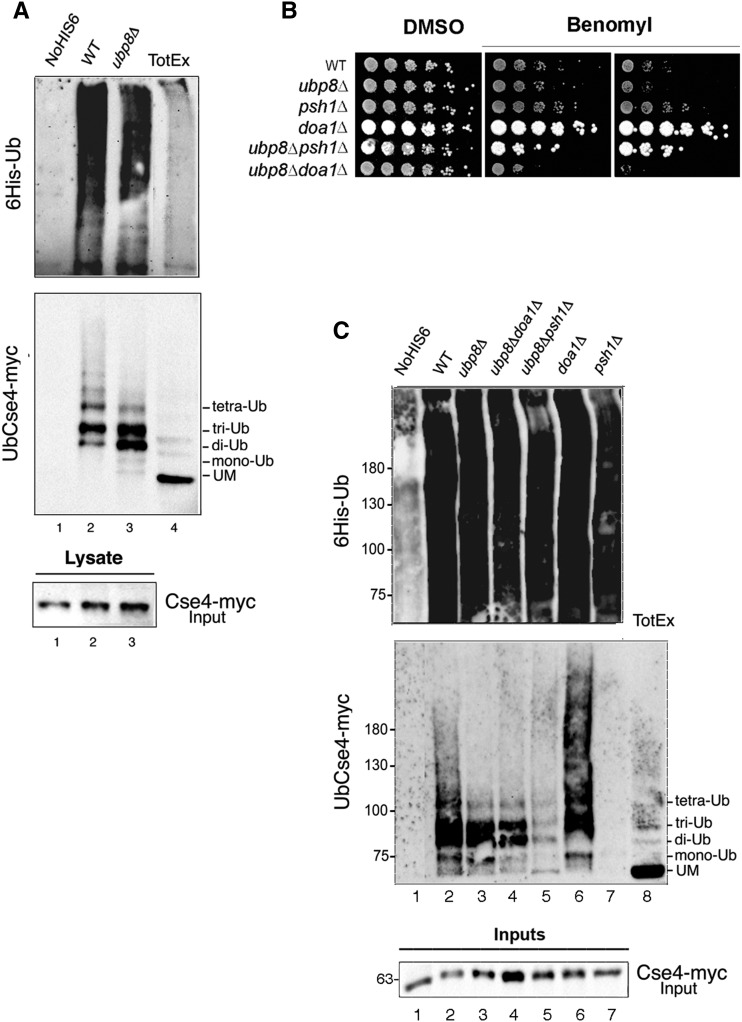
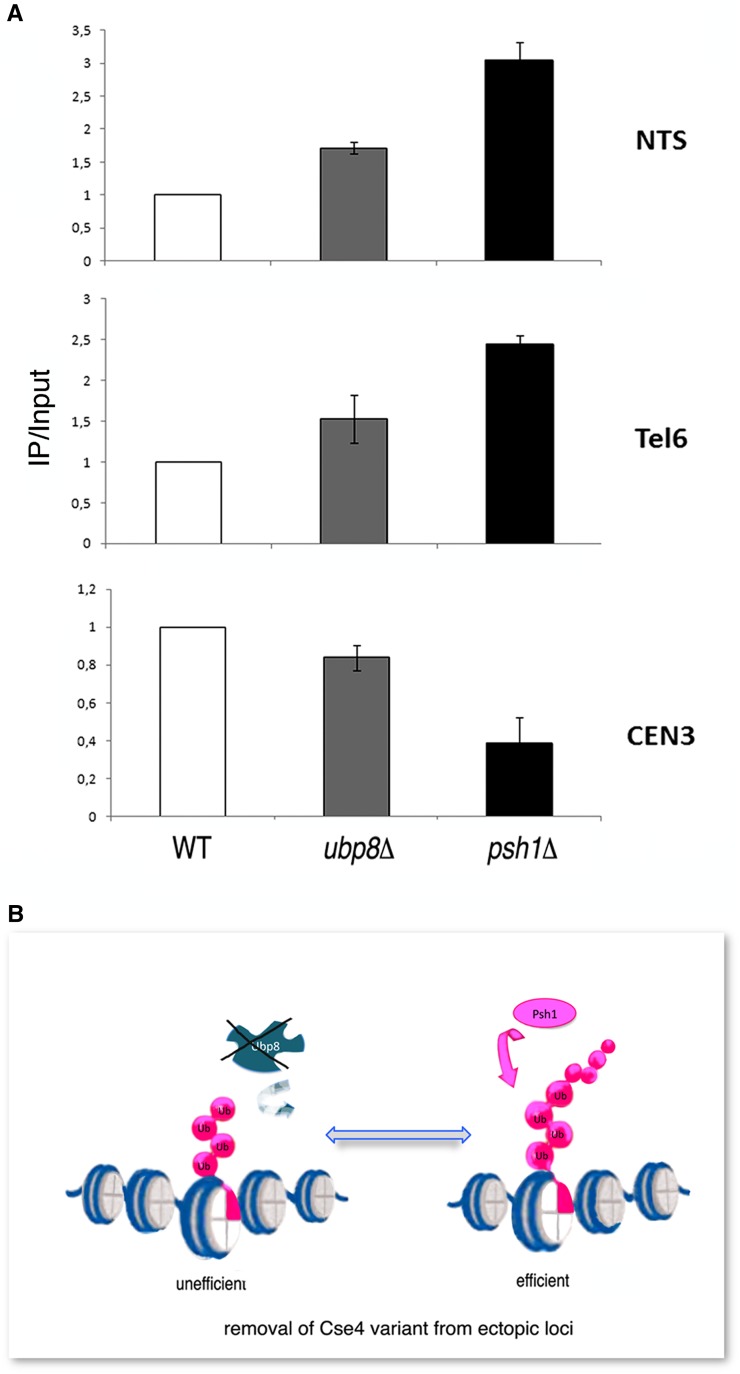
Aneuploidy, the unbalanced segregation of chromosomes during cell division, is recurrent in many tumors and the cause of birth defects and genetic diseases. Centromeric chromatin represents the chromosome attachment site to the mitotic spindle, marked by specialized nucleosomes containing a specific histone variant, CEN-H3/Cse4, in yeast. Mislocalization of Cse4 outside the centromere is deleterious and may cause aberrant chromosome behavior and mitotic loss. For this reason, ubiquitylation by the E3-ubiquitin ligase Psh1 and subsequent proteolysis tightly regulates its restricted localization. Among multiproteic machineries, the SAGA complex is not merely engaged in acetylation but also directly involved in deubiquitylation. In this study, we investigated the role of SAGA-DUB's Ubp8-driven deubiquitylation of the centromeric histone variant Cse4 in budding yeast. We found that Ubp8 works in concert with the E3-ubiquitin ligase Psh1, and that its loss causes defective deubiquitylation and the accumulation of a short ubiquitin oligomer on Cse4. We also show that lack of Ubp8 and defective deubiquitylation increase mitotic instability, cause faster Cse4 proteolysis and induce mislocalization of the centromeric histone outside the centromere. Our data provide evidence for a fundamental role of DUB-Ubp8 in deubiquitylation and the stability of the centromeric histone in budding yeast.
Keywords: DUB-Ubp8; SAGA complex; centromere; deubiquitylation; histone variant Cse4; mitotic stability.
Copyright © 2016 Canzonetta et al.
Figures
References
-
- Bernad R., Sánchez P., Losada A., 2009. Epigenetic specification of centromeres by CENP-A. Exp. Cell Res. 315(19): 3233–3241. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases