

Corosolic acid ameliorates acute inflammation through inhibition of IRAK-1 phosphorylation in macrophages
- PMID: 26615974
- PMCID: PMC5070707
- DOI: 10.5483/bmbrep.2016.49.5.241
Corosolic acid ameliorates acute inflammation through inhibition of IRAK-1 phosphorylation in macrophages
Abstract
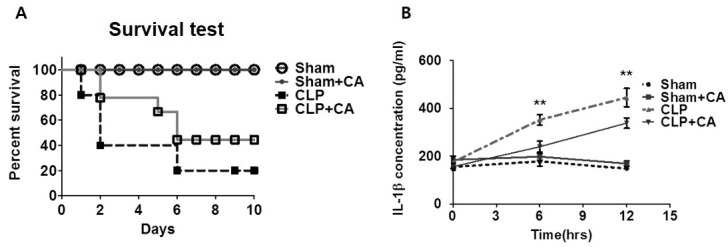
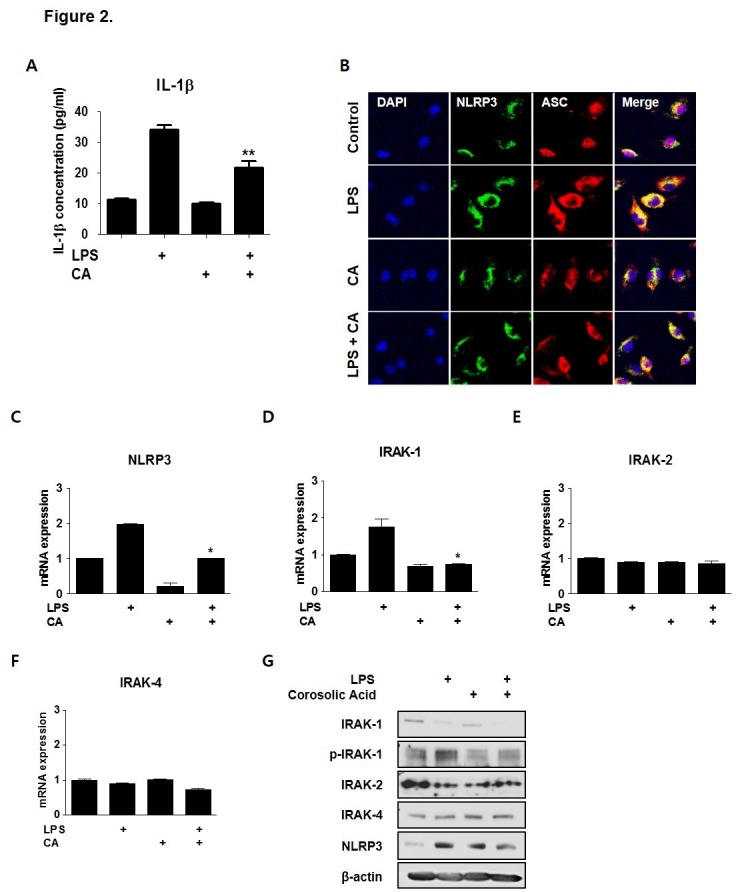
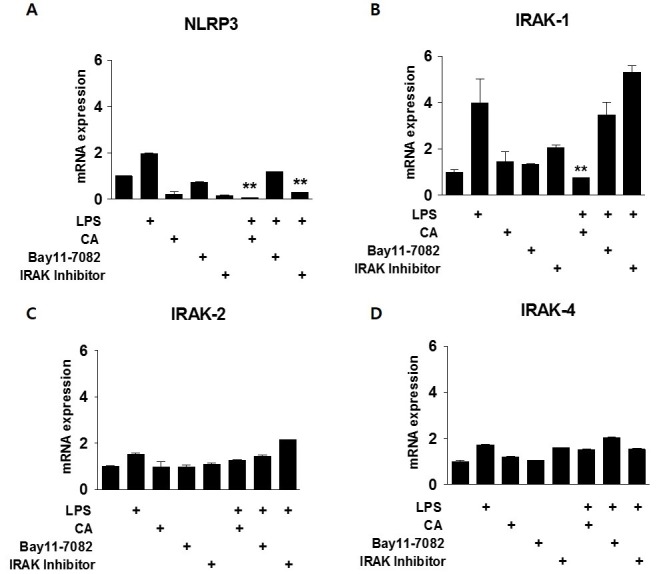
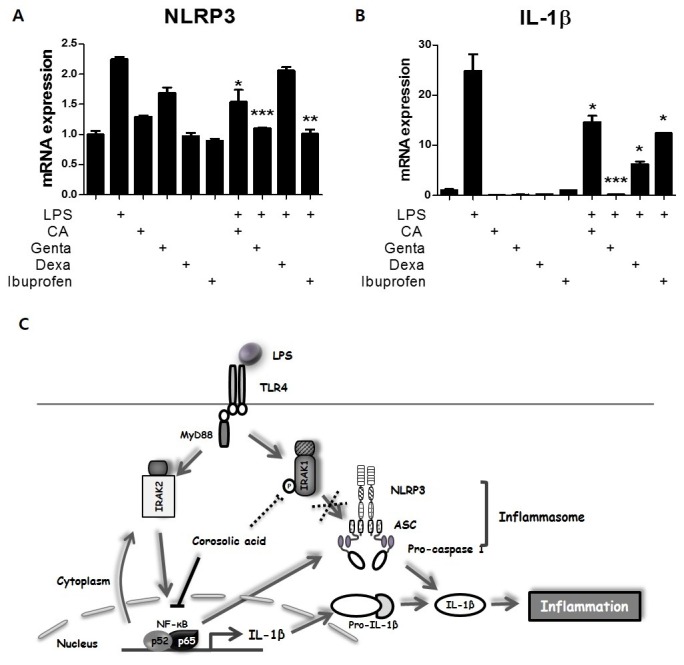
Corosolic acid (CA), a triterpenoid compound isolated from Lagerstroemia speciosa L. (Banaba) leaves, exerts anti-inflammatory effects by regulating phosphorylation of interleukin receptor-associated kinase (IRAK)-2 via the NF-κB cascade. However, the protective effect of CA against endotoxic shock has not been reported. LPS (200 ng/mL, 30 min) induced phosphorylation of IRAK-1 and treatment with CA (10 μM) significantly attenuated this effect. In addition, CA also reduced protein levels of NLRP3 and ASC which are the main components of the inflammasome in BMDMs. LPS-induced inflammasome assembly through activation of IRAK-1 was down-regulated by CA challenge. Treatment with Bay11-7082, an inhibitor of IκB-α, had no effect on CA-mediated inhibition of IRAK-1 activation, indicating that CA-mediated attenuation of IRAK-1 phosphorylation was independent of NF-κB signaling. These results demonstrate that CA ameliorates acute inflammation in mouse BMDMs and CA may be useful as a pharmacological agent to prevent acute inflammation. [BMB Reports 2016; 49(5): 276-281].
Figures
References
-
- Akira S. Toll-like receptors and innate immunity. Adv Immunol. (2001);78:1–56. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
