

Grid Cells and Place Cells: An Integrated View of their Navigational and Memory Function
- PMID: 26616686
- PMCID: PMC4679502
- DOI: 10.1016/j.tins.2015.10.004
Grid Cells and Place Cells: An Integrated View of their Navigational and Memory Function
Abstract
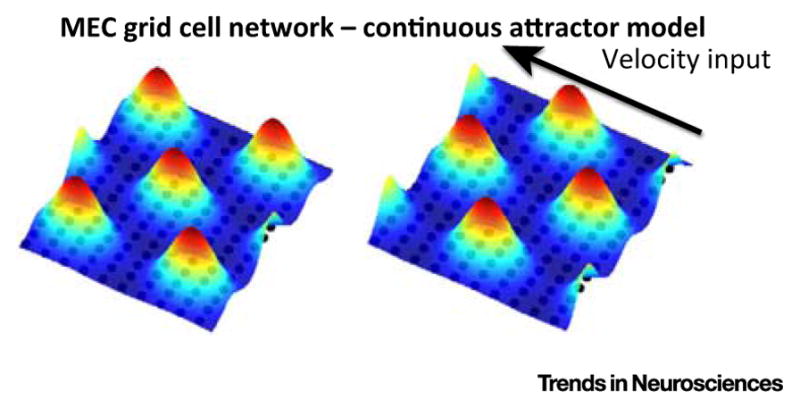
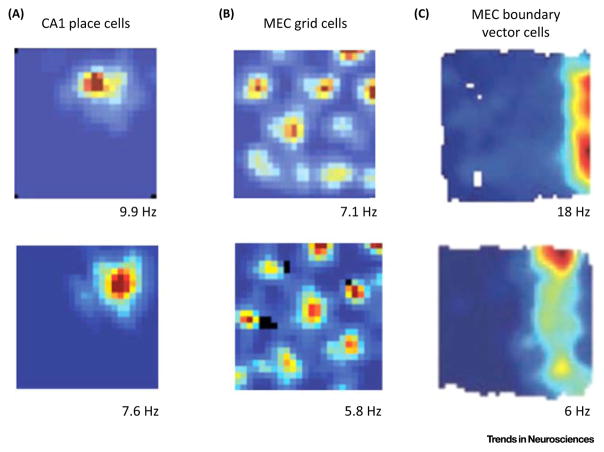
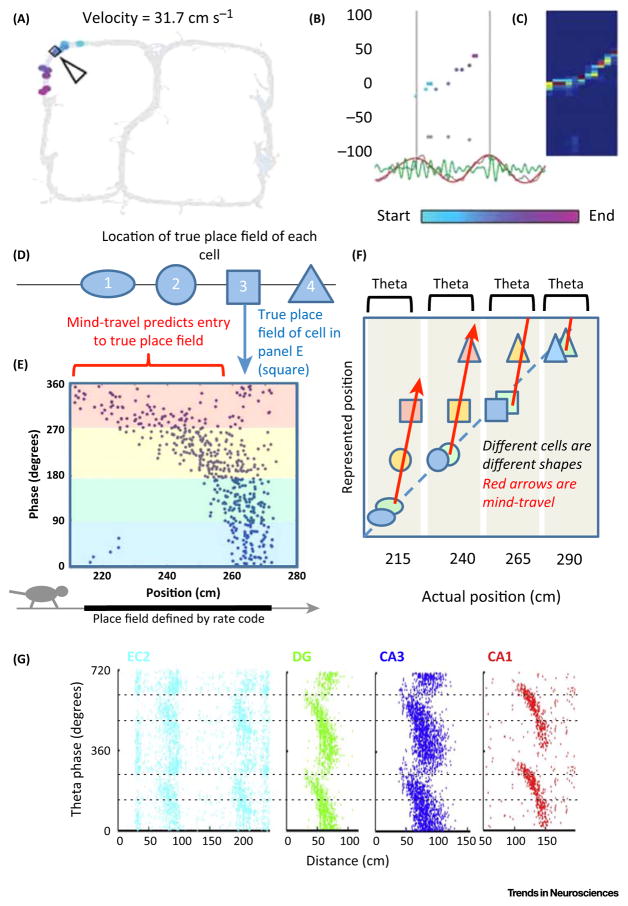
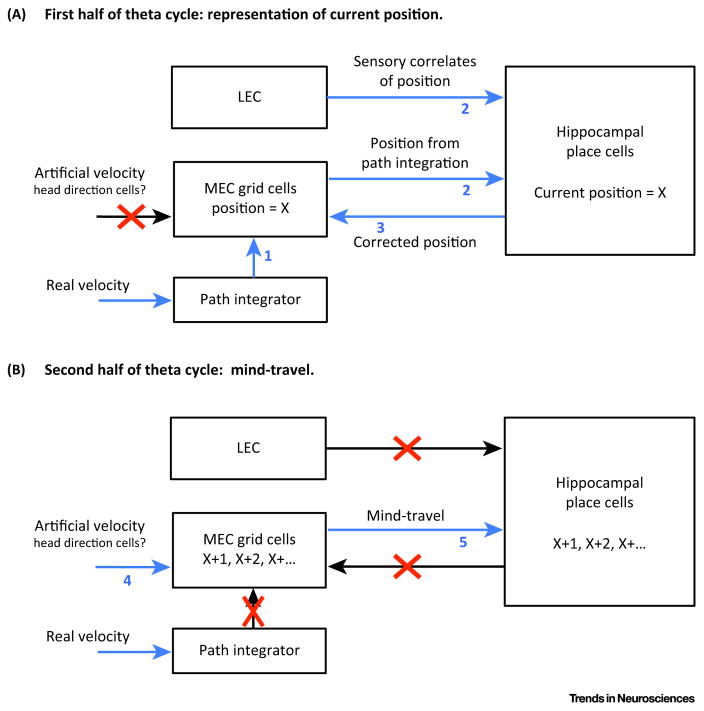
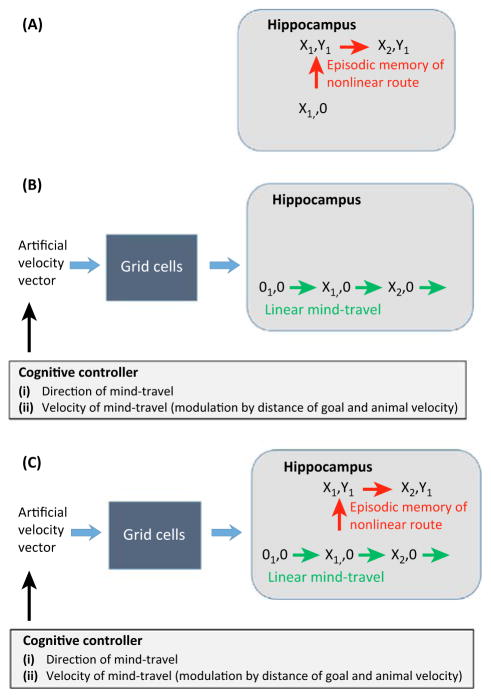
Much has been learned about the hippocampal/entorhinal system, but an overview of how its parts work in an integrated way is lacking. One question regards the function of entorhinal grid cells. We propose here that their fundamental function is to provide a coordinate system for producing mind-travel in the hippocampus, a process that accesses associations with upcoming positions. We further propose that mind-travel occurs during the second half of each theta cycle. By contrast, the first half of each theta cycle is devoted to computing current position using sensory information from the lateral entorhinal cortex (LEC) and path integration information from the medial entorhinal cortex (MEC). This model explains why MEC lesions can abolish hippocampal phase precession but not place fields.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- O’Keefe J. Place units in the hippocampus of the freely moving rat. Exp Neurol. 1976;51:78–109. - PubMed
-
- Hafting T, et al. Microstructure of a spatial map in the entorhinal cortex. Nature. 2005;436:801–806. - PubMed
-
- Sargolini F, et al. Conjunctive representation of position, direction, and velocity in entorhinal cortex. Science. 2006;312:758–762. - PubMed
-
- Taube JS. The head direction signal: origins and sensory-motor integration. Annu Rev Neurosci. 2007;30:181–207. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
