

Acetylation of glucokinase regulatory protein decreases glucose metabolism by suppressing glucokinase activity
- PMID: 26620281
- PMCID: PMC4664969
- DOI: 10.1038/srep17395
Acetylation of glucokinase regulatory protein decreases glucose metabolism by suppressing glucokinase activity
Abstract
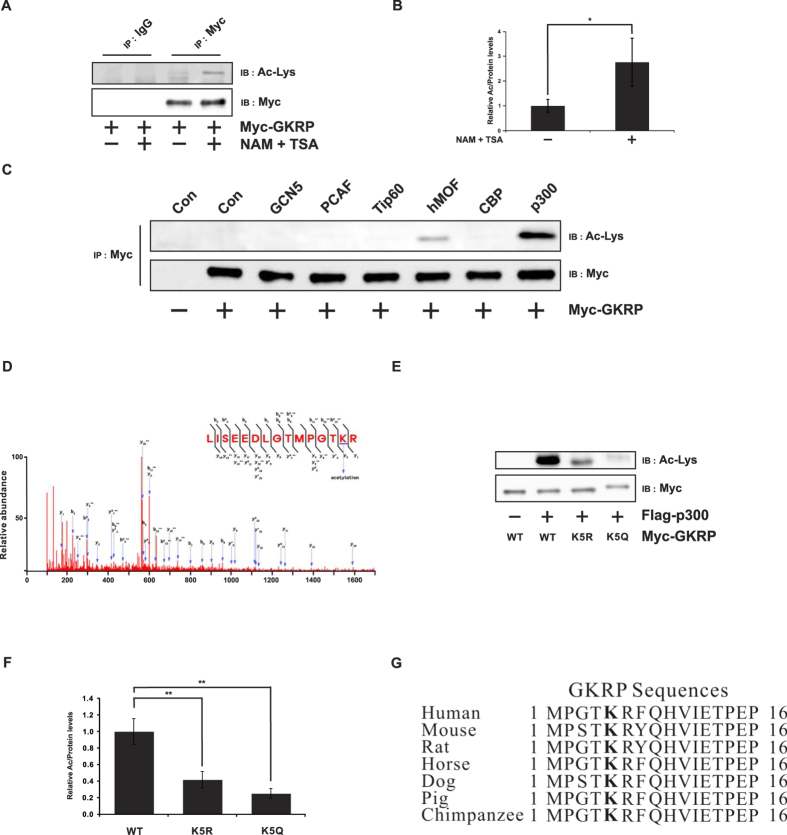
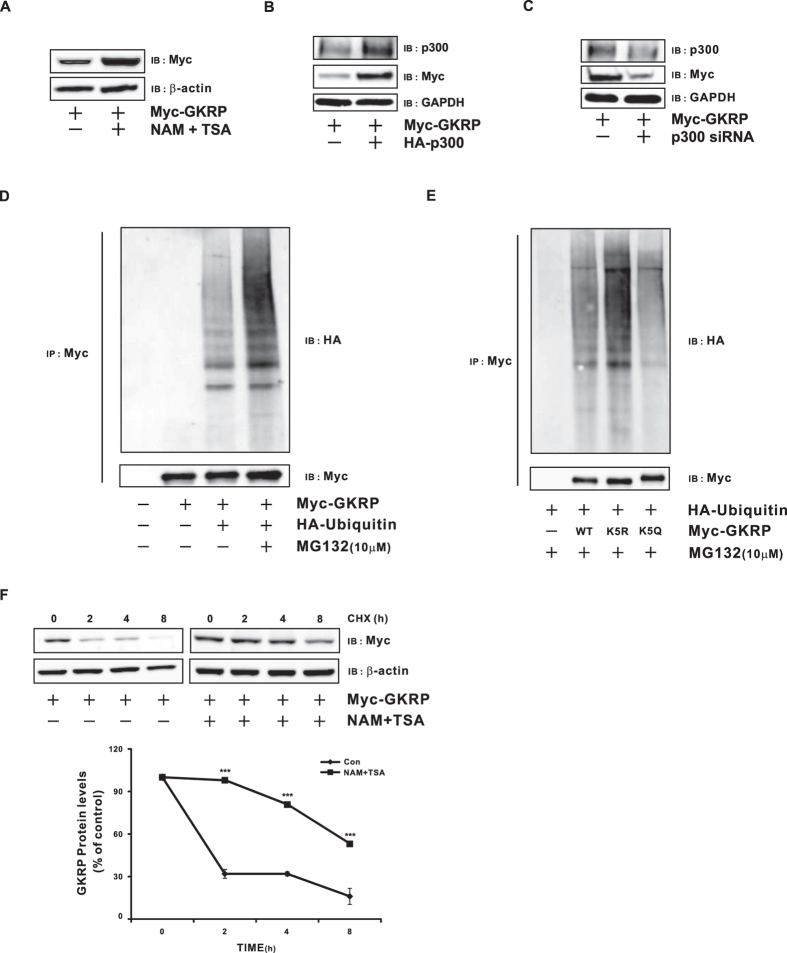
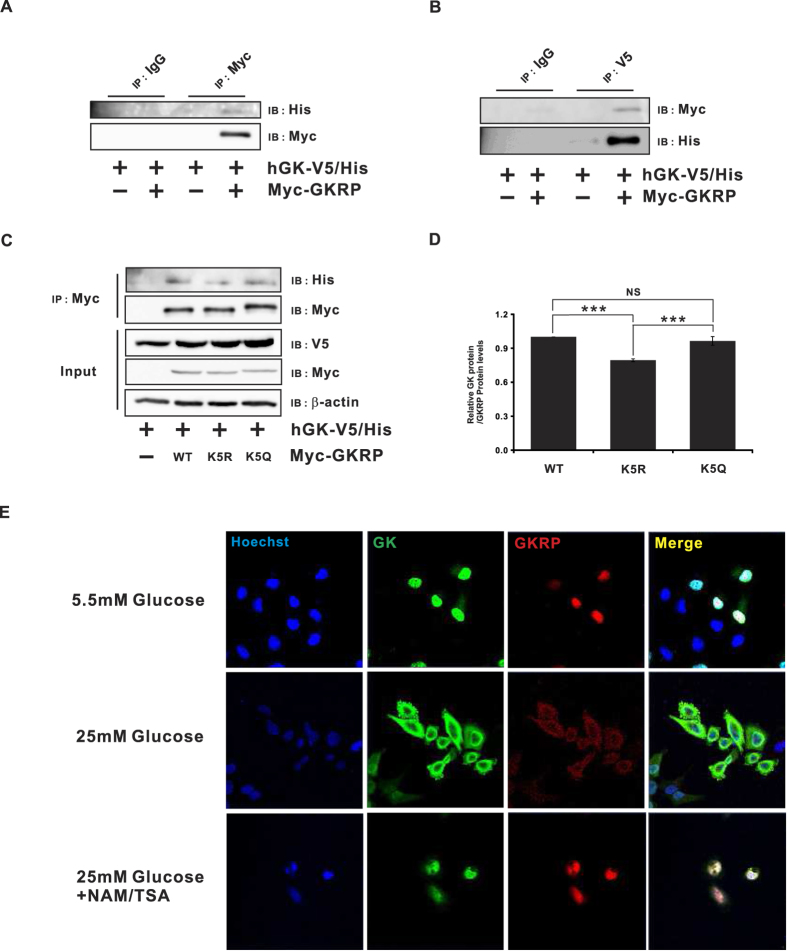
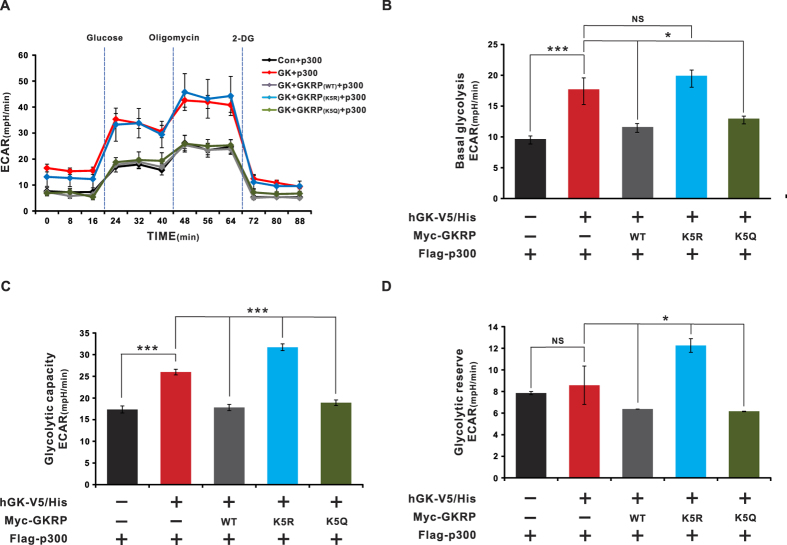
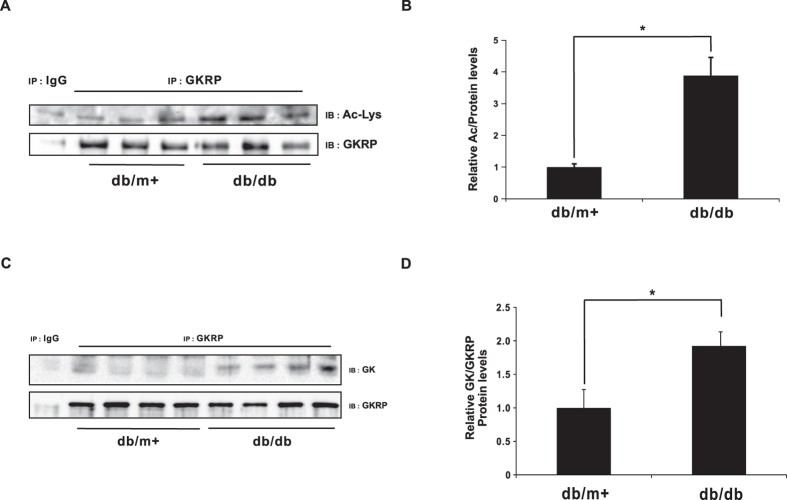
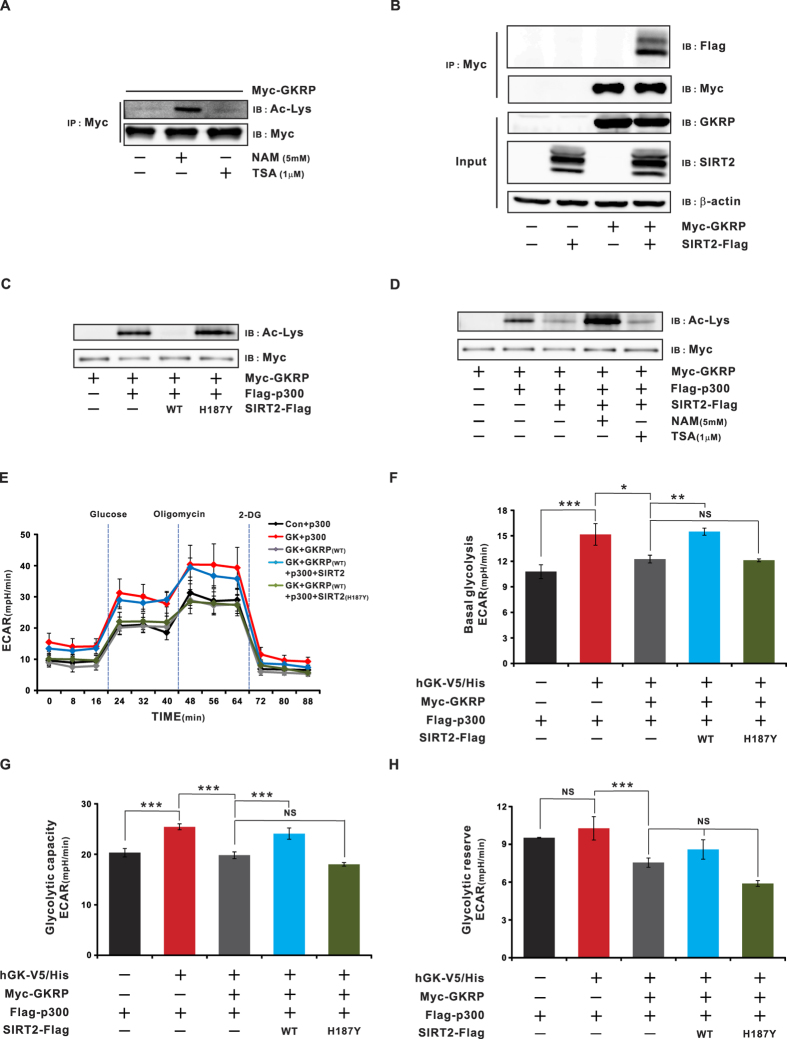
Glucokinase (GK), mainly expressed in the liver and pancreatic β-cells, is critical for maintaining glucose homeostasis. GK expression and kinase activity, respectively, are both modulated at the transcriptional and post-translational levels. Post-translationally, GK is regulated by binding the glucokinase regulatory protein (GKRP), resulting in GK retention in the nucleus and its inability to participate in cytosolic glycolysis. Although hepatic GKRP is known to be regulated by allosteric mechanisms, the precise details of modulation of GKRP activity, by post-translational modification, are not well known. Here, we demonstrate that GKRP is acetylated at Lys5 by the acetyltransferase p300. Acetylated GKRP is resistant to degradation by the ubiquitin-dependent proteasome pathway, suggesting that acetylation increases GKRP stability and binding to GK, further inhibiting GK nuclear export. Deacetylation of GKRP is effected by the NAD(+)-dependent, class III histone deacetylase SIRT2, which is inhibited by nicotinamide. Moreover, the livers of db/db obese, diabetic mice also show elevated GKRP acetylation, suggesting a broader, critical role in regulating blood glucose. Given that acetylated GKRP may affiliate with type-2 diabetes mellitus (T2DM), understanding the mechanism of GKRP acetylation in the liver could reveal novel targets within the GK-GKRP pathway, for treating T2DM and other metabolic pathologies.
Figures
References
-
- Mokdad A. H. et al. Prevalence of obesity, diabetes, and obesity-related health risk factors, 2001. JAMA 289, 76–79 (2003). - PubMed
-
- Consoli A. Role of liver in pathophysiology of NIDDM. Diabetes Care 15, 430–441 (1992). - PubMed
-
- Matschinsky F. M. Glucokinase as glucose sensor and metabolic signal generator in pancreatic beta-cells and hepatocytes. Diabetes 39, 647–652 (1990). - PubMed
-
- Nordlie R. C., Foster J. D. & Lange A. J. Regulation of glucose production by the liver. Annu Rev Nutr 19, 379–406 (1999). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
