

Volume-regulated anion channel--a frenemy within the brain
- PMID: 26620797
- PMCID: PMC4752865
- DOI: 10.1007/s00424-015-1765-6
Volume-regulated anion channel--a frenemy within the brain
Abstract
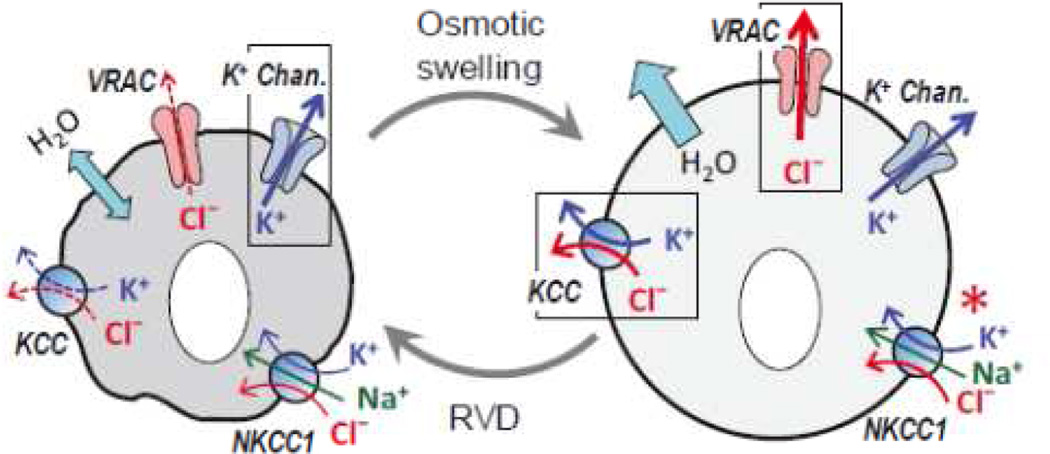
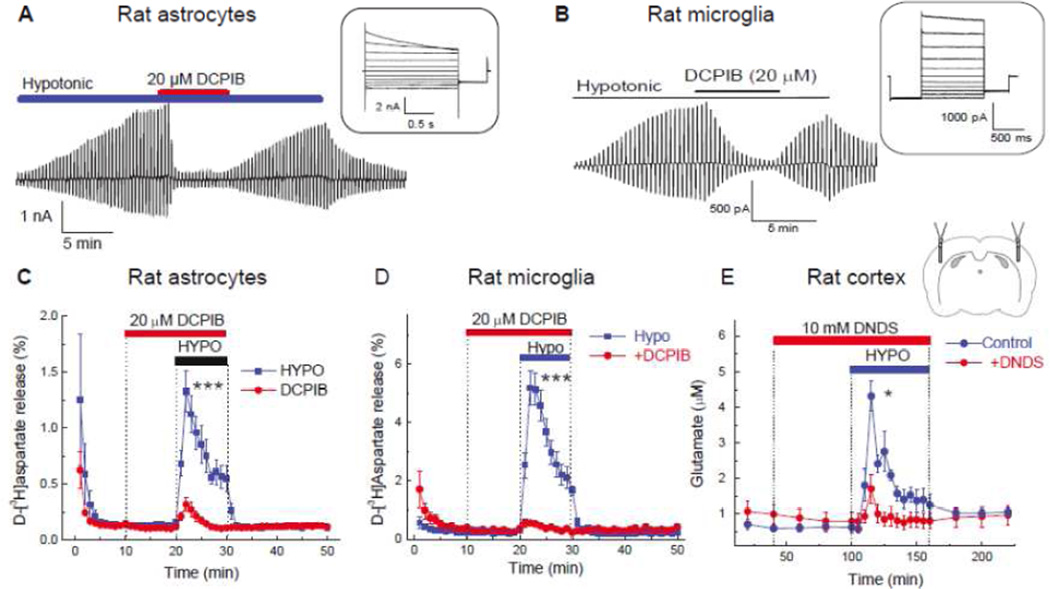
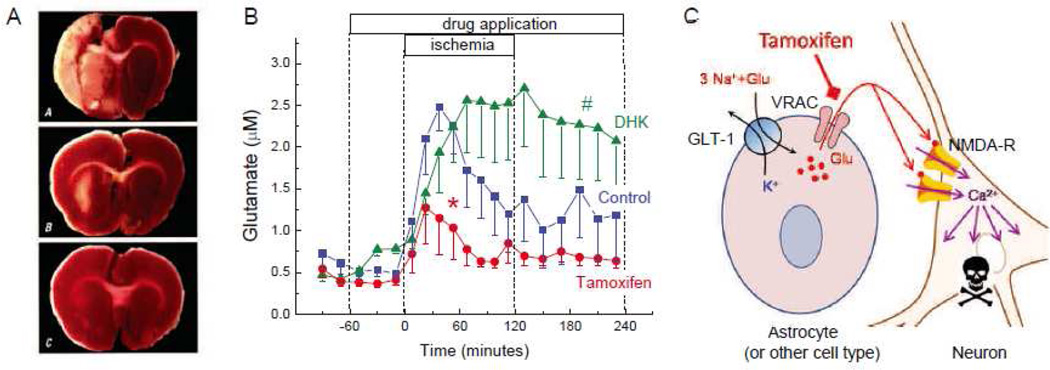
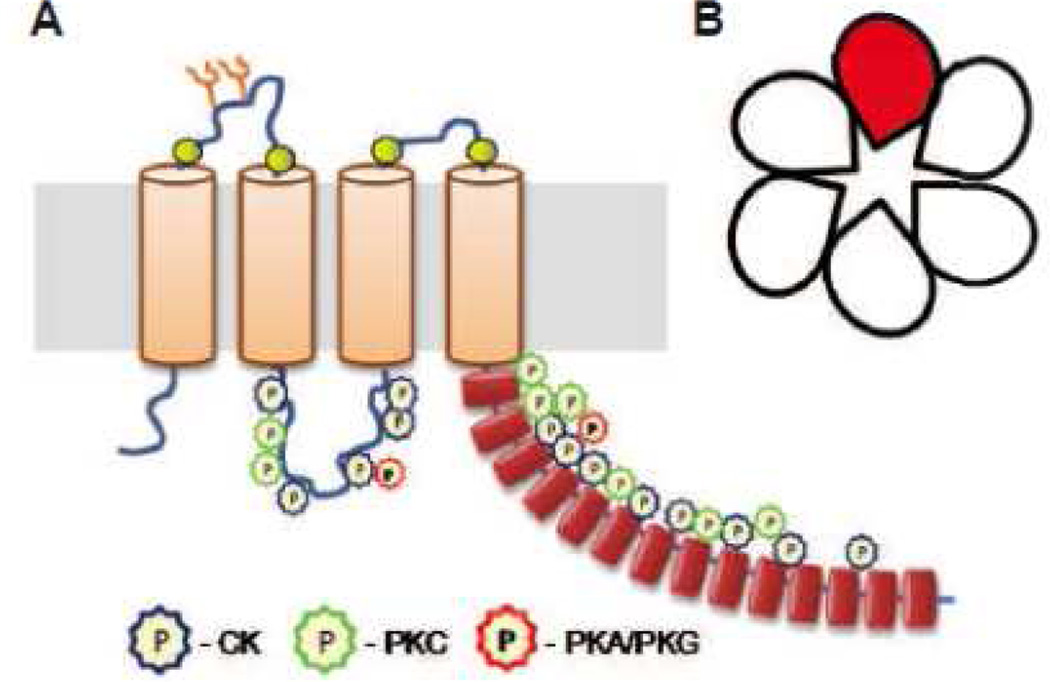
The volume-regulated anion channel (VRAC) is a ubiquitously expressed yet highly enigmatic member of the superfamily of chloride/anion channels. It is activated by cellular swelling and mediates regulatory cell volume decrease in a majority of vertebrate cells, including those in the central nervous system (CNS). In the brain, besides its crucial role in cellular volume regulation, VRAC is thought to play a part in cell proliferation, apoptosis, migration, and release of physiologically active molecules. Although these roles are not exclusive to the CNS, the relative significance of VRAC in the brain is amplified by several unique aspects of its physiology. One important example is the contribution of VRAC to the release of the excitatory amino acid neurotransmitters glutamate and aspartate. This latter process is thought to have impact on both normal brain functioning (such as astrocyte-neuron signaling) and neuropathology (via promoting the excitotoxic death of neuronal cells in stroke and traumatic brain injury). In spite of much work in the field, the molecular nature of VRAC remained unknown until less than 2 years ago. Two pioneer publications identified VRAC as the heterohexamer formed by the leucine-rich repeat-containing 8 (LRRC8) proteins. These findings galvanized the field and are likely to result in dramatic revisions to our understanding of the place and role of VRAC in the brain, as well as other organs and tissues. The present review briefly recapitulates critical findings in the CNS and focuses on anticipated impact on the LRRC8 discovery on further progress in neuroscience research.
Keywords: Astrocytes; Cell volume regulation; Central nervous system; LRRC8; Neurons; VRAC.
Figures
Similar articles
-
Molecular Biology and Physiology of Volume-Regulated Anion Channel (VRAC).Curr Top Membr. 2018;81:177-203. doi: 10.1016/bs.ctm.2018.07.005. Epub 2018 Aug 14. Curr Top Membr. 2018. PMID: 30243432 Free PMC article. Review.
-
Biophysics and Physiology of the Volume-Regulated Anion Channel (VRAC)/Volume-Sensitive Outwardly Rectifying Anion Channel (VSOR).Pflugers Arch. 2016 Mar;468(3):371-83. doi: 10.1007/s00424-015-1781-6. Epub 2016 Jan 6. Pflugers Arch. 2016. PMID: 26739710 Review.
-
The volume-regulated anion channel is formed by LRRC8 heteromers – molecular identification and roles in membrane transport and physiology.Biol Chem. 2015 Sep;396(9-10):975-90. doi: 10.1515/hsz-2015-0127. Biol Chem. 2015. PMID: 25868000 Review.
-
Molecular composition and heterogeneity of the LRRC8-containing swelling-activated osmolyte channels in primary rat astrocytes.J Physiol. 2017 Nov 15;595(22):6939-6951. doi: 10.1113/JP275053. Epub 2017 Sep 12. J Physiol. 2017. PMID: 28833202 Free PMC article.
-
The identification of a volume-regulated anion channel: an amazing Odyssey.Acta Physiol (Oxf). 2015 Apr;213(4):868-81. doi: 10.1111/apha.12450. Epub 2015 Jan 28. Acta Physiol (Oxf). 2015. PMID: 25565132 Review.
Cited by
-
Taurine Supplementation as a Neuroprotective Strategy upon Brain Dysfunction in Metabolic Syndrome and Diabetes.Nutrients. 2022 Mar 18;14(6):1292. doi: 10.3390/nu14061292. Nutrients. 2022. PMID: 35334949 Free PMC article. Review.
-
Binding of the protein ICln to α-integrin contributes to the activation of IClswell current.Sci Rep. 2019 Aug 21;9(1):12195. doi: 10.1038/s41598-019-48496-4. Sci Rep. 2019. PMID: 31434921 Free PMC article.
-
The LRRC8/VRAC anion channel facilitates myogenic differentiation of murine myoblasts by promoting membrane hyperpolarization.J Biol Chem. 2019 Sep 27;294(39):14279-14288. doi: 10.1074/jbc.RA119.008840. Epub 2019 Aug 6. J Biol Chem. 2019. PMID: 31387946 Free PMC article.
-
Neurodegeneration Upon Dysfunction of Endosomal/Lysosomal CLC Chloride Transporters.Front Cell Dev Biol. 2021 Feb 23;9:639231. doi: 10.3389/fcell.2021.639231. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33708769 Free PMC article. Review.
-
Gene Network Dysregulation in the Trigeminal Ganglia and Nucleus Accumbens of a Model of Chronic Migraine-Associated Hyperalgesia.Front Syst Neurosci. 2018 Dec 18;12:63. doi: 10.3389/fnsys.2018.00063. eCollection 2018. Front Syst Neurosci. 2018. PMID: 30618656 Free PMC article.
References
-
- Abascal F, Zardoya R. LRRC8 proteins share a common ancestor with pannexins, and may form hexameric channels involved in cell-cell communication. Bioessays. 2012;34(7):551–560. - PubMed
-
- Adrogue HJ, Madias NE. Hyponatremia. N Engl J Med. 2000;342(21):1581–1589. - PubMed
-
- Aitken PG, Borgdorff AJ, Juta AJ, Kiehart DP, Somjen GG, Wadman WJ. Volume changes induced by osmotic stress in freshly isolated rat hippocampal neurons. Pflugers Arch. 1998;436(6):991–998. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
