

Peripheral Circadian Clocks Mediate Dietary Restriction-Dependent Changes in Lifespan and Fat Metabolism in Drosophila
- PMID: 26626459
- PMCID: PMC4715572
- DOI: 10.1016/j.cmet.2015.10.014
Peripheral Circadian Clocks Mediate Dietary Restriction-Dependent Changes in Lifespan and Fat Metabolism in Drosophila
Abstract
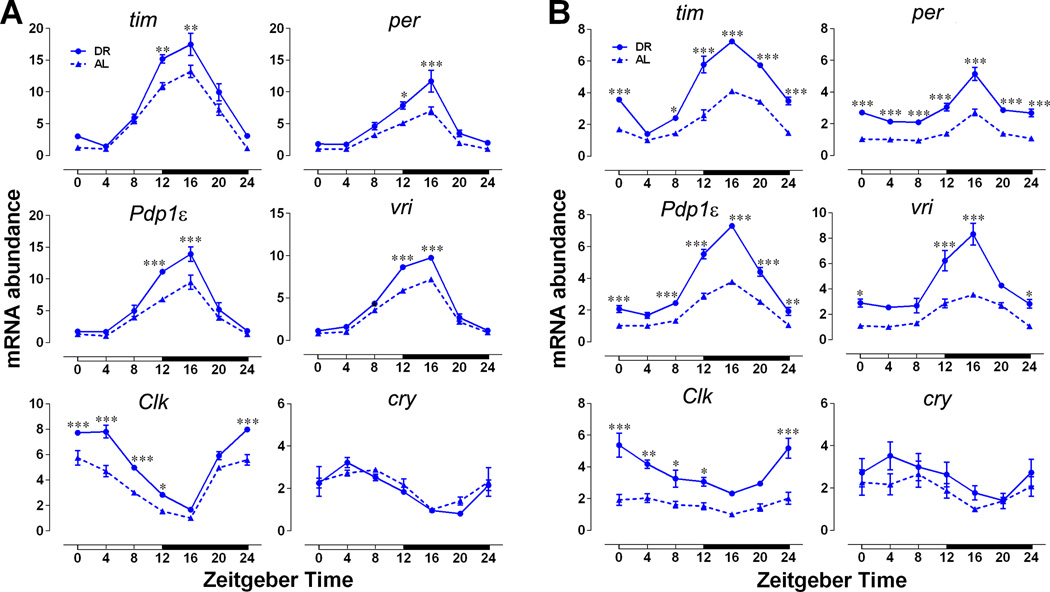
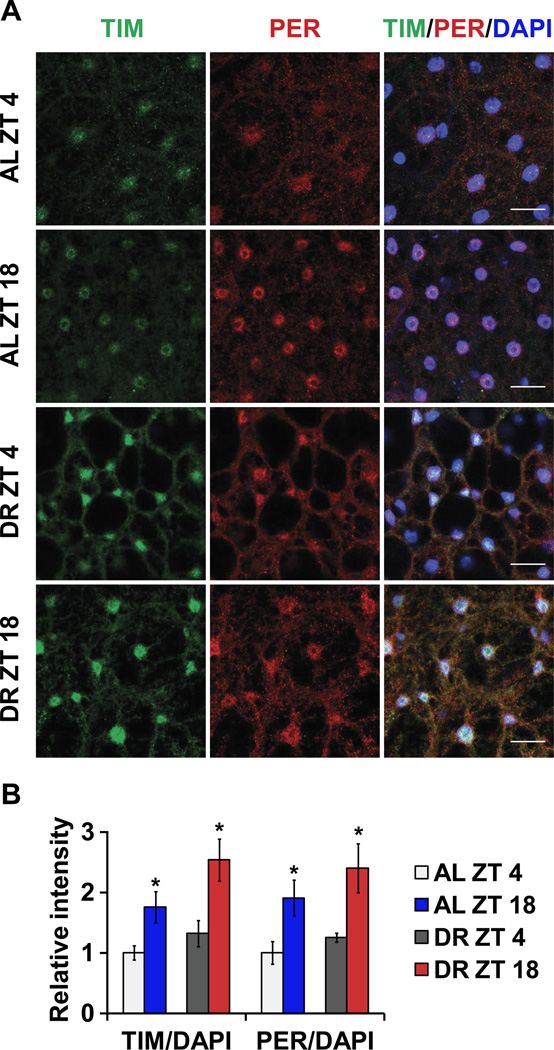
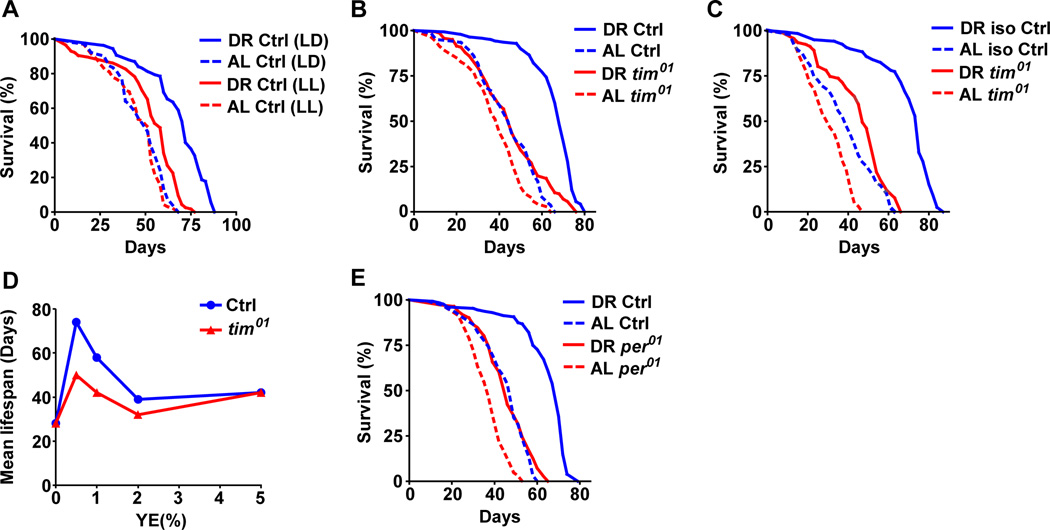
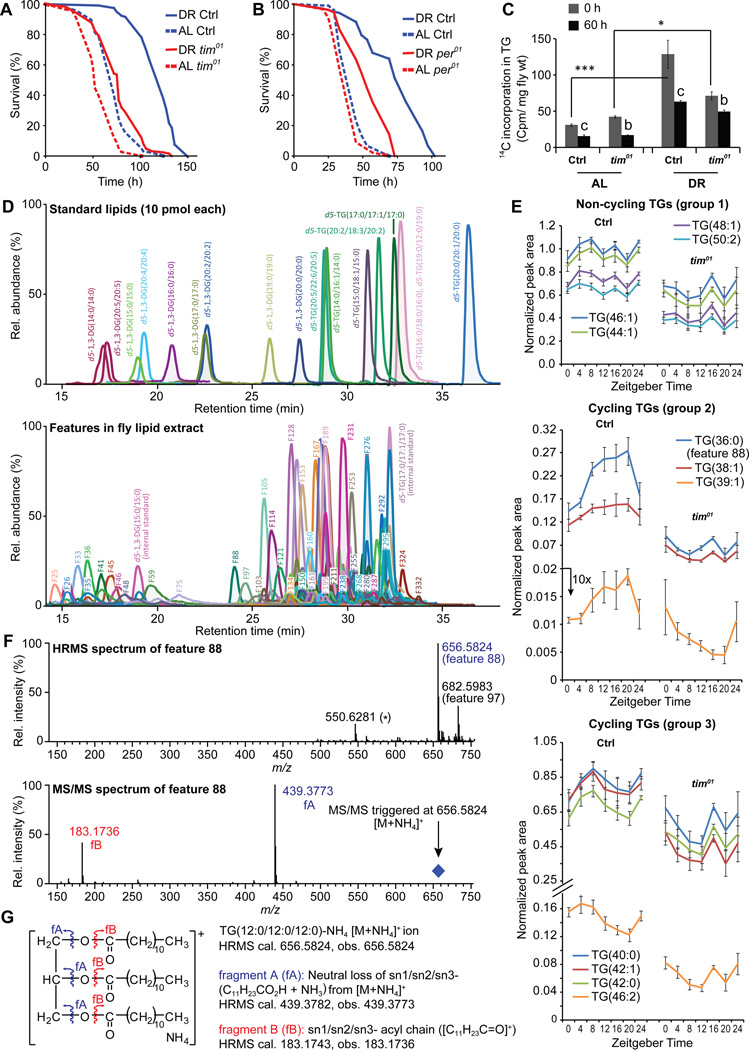
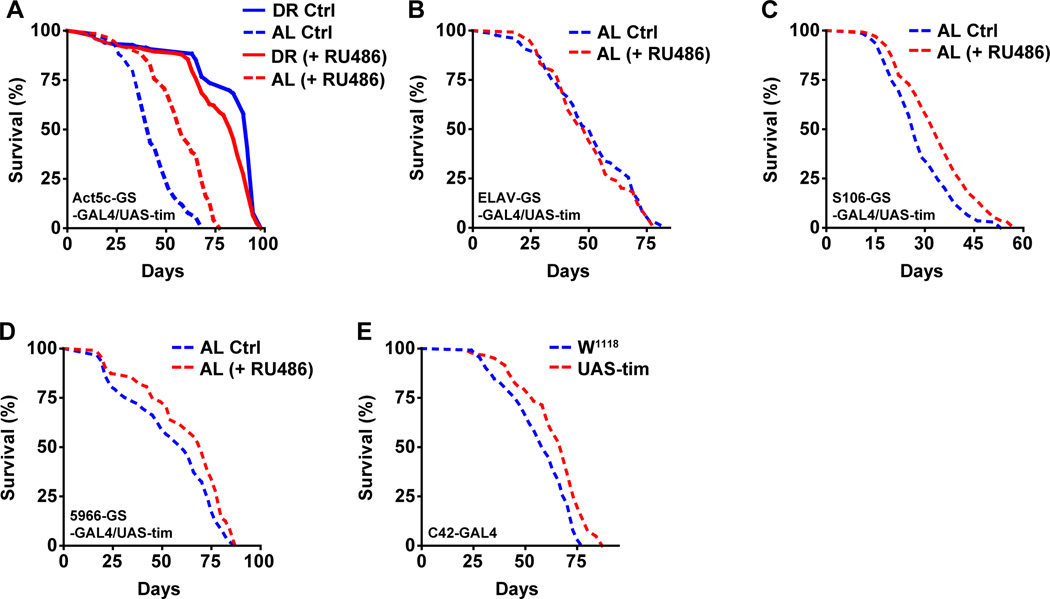
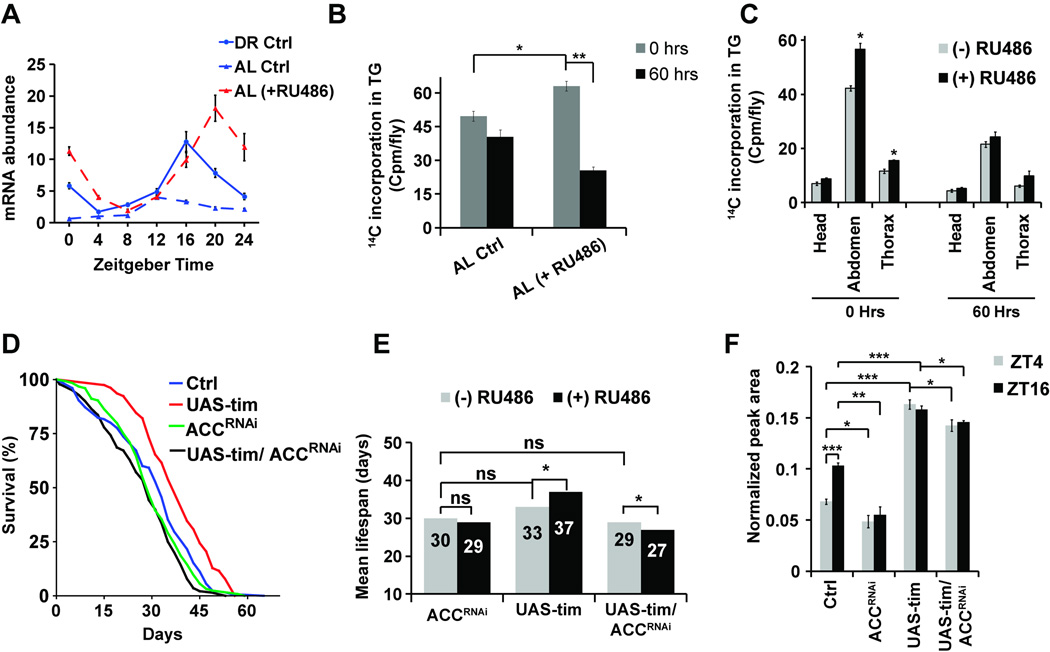
Endogenous circadian clocks orchestrate several metabolic and signaling pathways that are known to modulate lifespan, suggesting clocks as potential targets for manipulation of metabolism and lifespan. We report here that the core circadian clock genes, timeless (tim) and period (per), are required for the metabolic and lifespan responses to DR in Drosophila. Consistent with the involvement of a circadian mechanism, DR enhances the amplitude of cycling of most circadian clock genes, including tim, in peripheral tissues. Mass-spectrometry-based lipidomic analysis suggests a role of tim in cycling of specific medium chain triglycerides under DR. Furthermore, overexpression of tim in peripheral tissues improves its oscillatory amplitude and extends lifespan under ad libitum conditions. Importantly, effects of tim on lifespan appear to be mediated through enhanced fat turnover. These findings identify a critical role for specific clock genes in modulating the effects of nutrient manipulation on fat metabolism and aging.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Dietary Restriction Extends the Lifespan of Circadian Mutants tim and per.Cell Metab. 2016 Dec 13;24(6):763-764. doi: 10.1016/j.cmet.2016.11.002. Epub 2016 Dec 1. Cell Metab. 2016. PMID: 27916531 Free PMC article. No abstract available.
References
-
- Asai M, Yoshinobu Y, Kaneko S, Mori A, Nikaido T, Moriya T, Akiyama M, Shibata S. Circadian profile of Per gene mRNA expression in the suprachiasmatic nucleus, paraventricular nucleus, and pineal body of aged rats. J. Neuro. Res. 2001;66:1133–1139. - PubMed
-
- Bishop NA, Guarente L. Genetic links between diet and lifespan: shared mechanisms from yeast to humans. Nat. Rev. Genet. 2007;8:835–844. - PubMed
-
- Bordone L, Guarente L. Calorie restriction, SIRT1 and metabolism: understanding longevity. Nat. Rev. Mol. Cell Biol. 2005;6:298–305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
