

Inhibition of poly(ADP-ribose)polymerase-1 and DNA repair by uranium
- PMID: 26627003
- PMCID: PMC4718819
- DOI: 10.1016/j.taap.2015.11.017
Inhibition of poly(ADP-ribose)polymerase-1 and DNA repair by uranium
Abstract
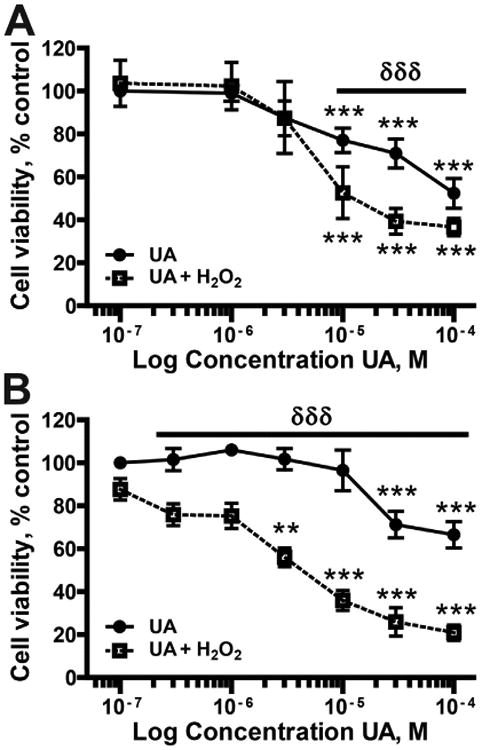
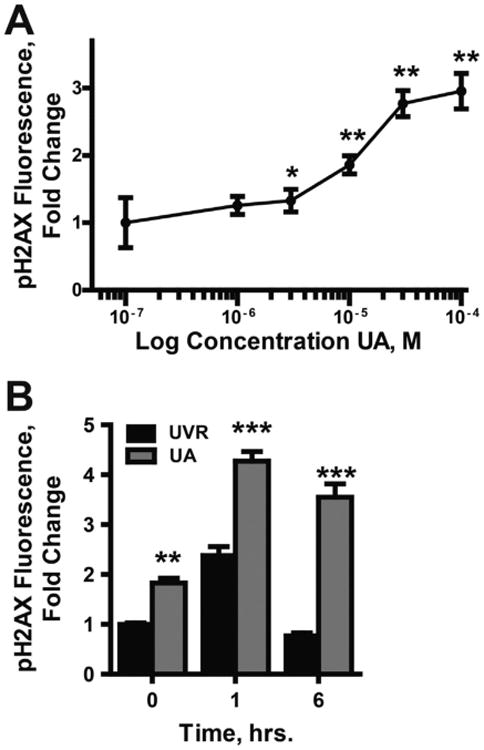
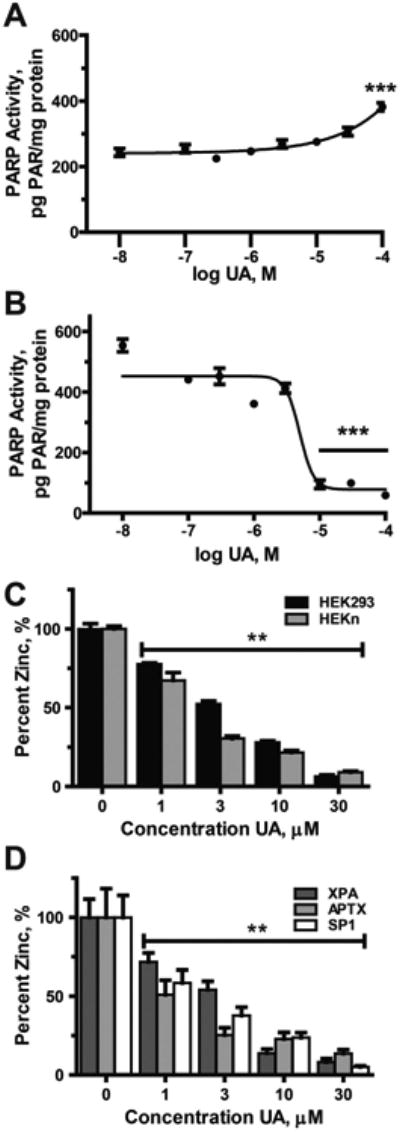
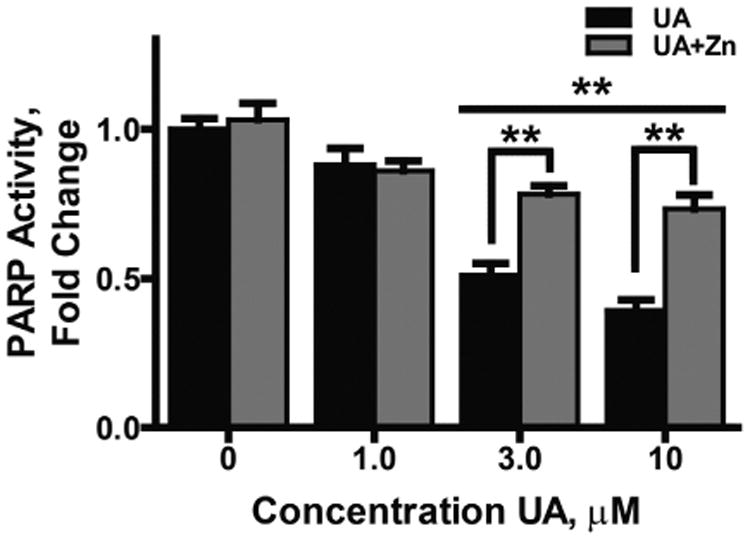
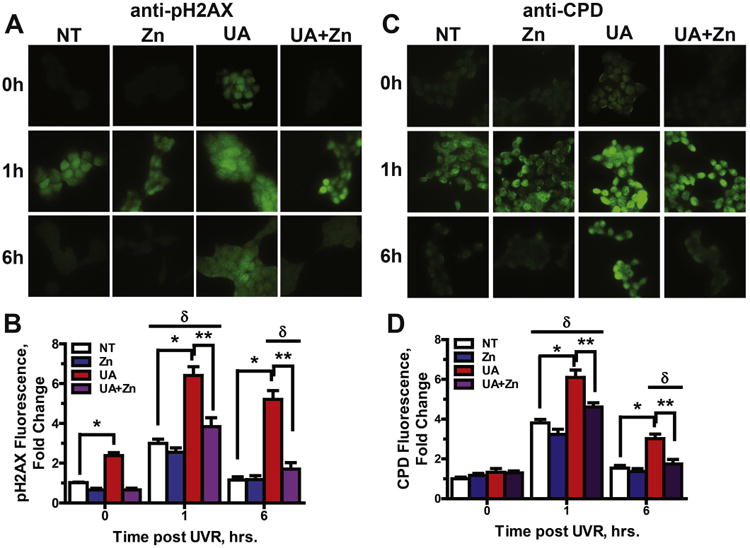
Uranium has radiological and non-radiological effects within biological systems and there is increasing evidence for genotoxic and carcinogenic properties attributable to uranium through its heavy metal properties. In this study, we report that low concentrations of uranium (as uranyl acetate; <10 μM) is not cytotoxic to human embryonic kidney cells or normal human keratinocytes; however, uranium exacerbates DNA damage and cytotoxicity induced by hydrogen peroxide, suggesting that uranium may inhibit DNA repair processes. Concentrations of uranyl acetate in the low micromolar range inhibited the zinc finger DNA repair protein poly(ADP-ribose) polymerase (PARP)-1 and caused zinc loss from PARP-1 protein. Uranyl acetate exposure also led to zinc loss from the zinc finger DNA repair proteins Xeroderma Pigmentosum, Complementation Group A (XPA) and aprataxin (APTX). In keeping with the observed inhibition of zinc finger function of DNA repair proteins, exposure to uranyl acetate enhanced retention of induced DNA damage. Co-incubation of uranyl acetate with zinc largely overcame the impact of uranium on PARP-1 activity and DNA damage. These findings present evidence that low concentrations of uranium can inhibit DNA repair through disruption of zinc finger domains of specific target DNA repair proteins. This may provide a mechanistic basis to account for the published observations that uranium exposure is associated with DNA repair deficiency in exposed human populations.
Keywords: DNA damage; DNA repair; Poly(ADP-ribose) polymerase-1 (PARP-1); Uranium; Zinc finger.
Copyright © 2015 Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures
References
-
- Au WW, McConnell MA, Wilkinson GS, Ramanujam VM, Alcock N. Population monitoring: experience with residents exposed to uranium mining/milling waste. Mutat Res. 1998;405:237–245. - PubMed
-
- Au WW, Giri AK, Ruchirawat M. Challenge assay: a functional biomarker for exposure-induced DNA repair deficiency and for risk of cancer. Int J Hyg Environ Health. 2010;213:32–39. - PubMed
-
- Bal W, Protas AM, Kasprzak KS. Genotoxicity of metal ions: chemical insights. Met Ions Life Sci. 2011;8:319–373. - PubMed
-
- Beyersmann D, Hartwig A. Carcinogenic metal compounds: recent insight into molecular and cellular mechanisms. Arch Toxicol. 2008;82:493–512. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
