

A quantitative acoustic analysis of the vocal repertoire of the common marmoset (Callithrix jacchus)
- PMID: 26627765
- PMCID: PMC4644241
- DOI: 10.1121/1.4934268
A quantitative acoustic analysis of the vocal repertoire of the common marmoset (Callithrix jacchus)
Abstract
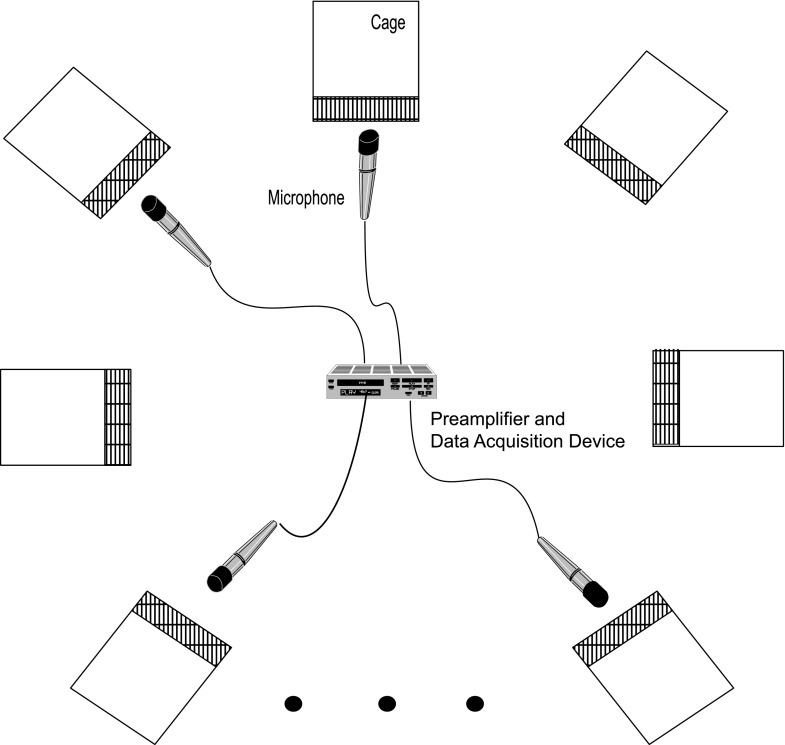
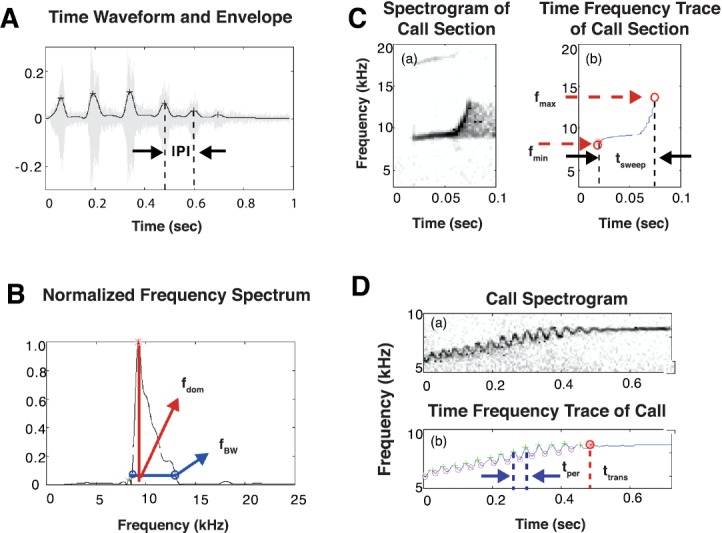
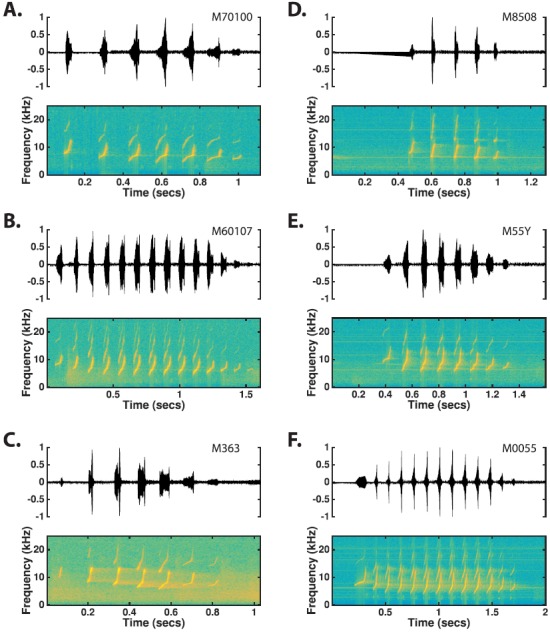
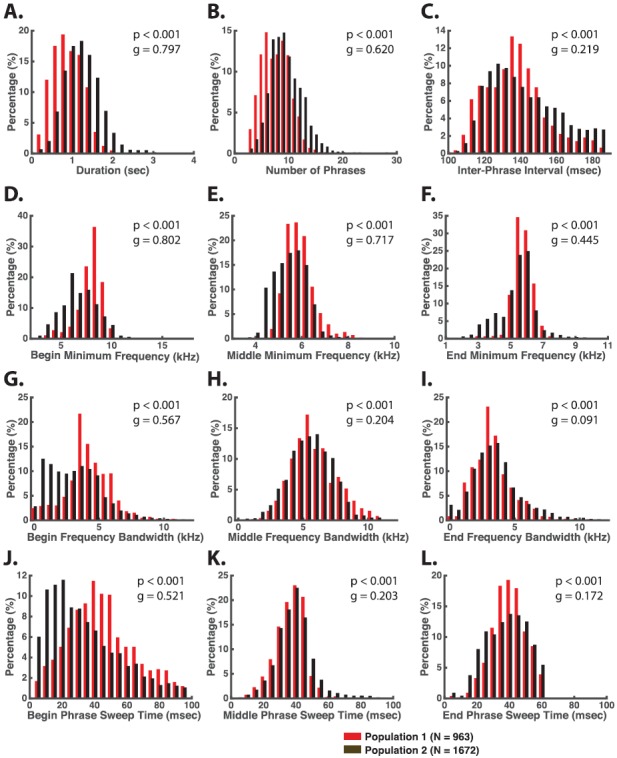
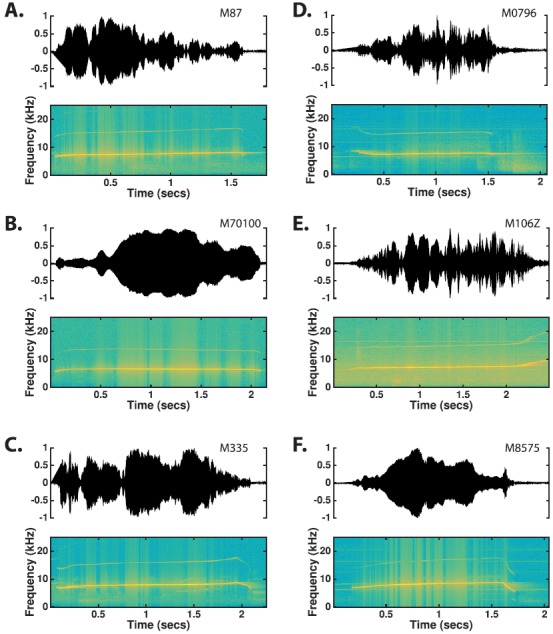
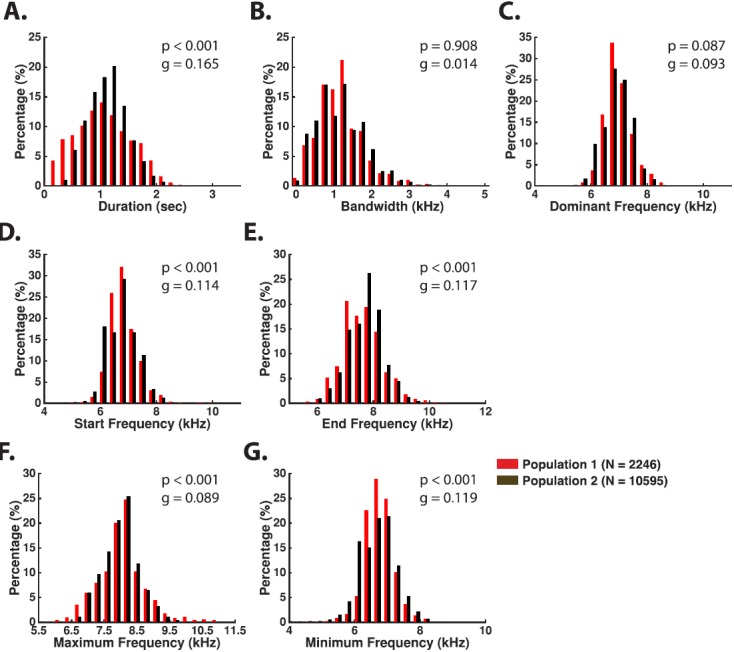
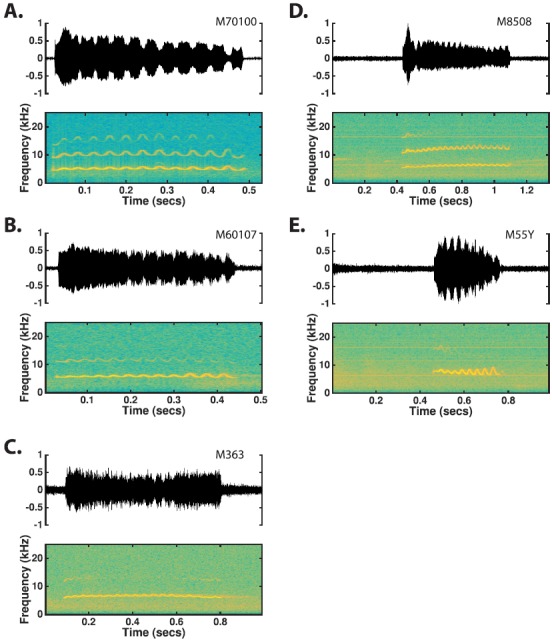
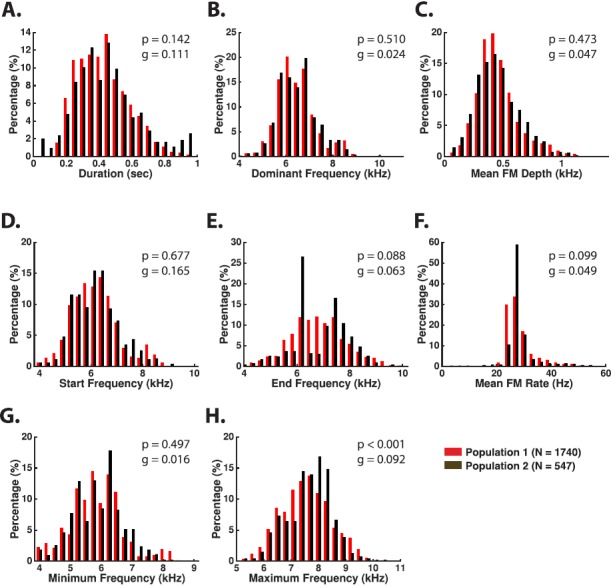
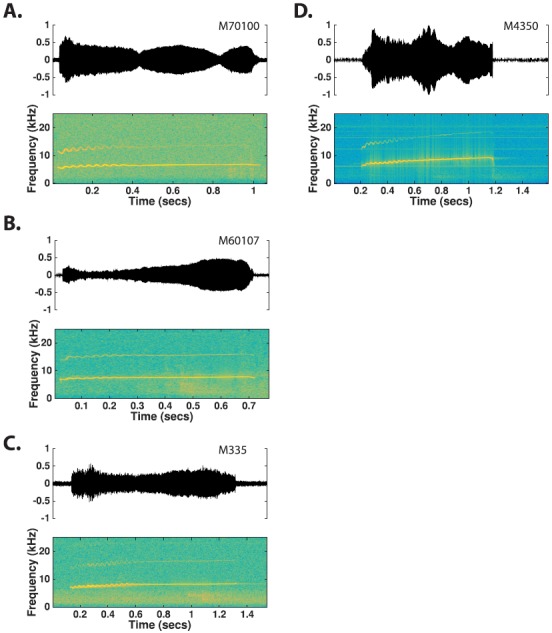
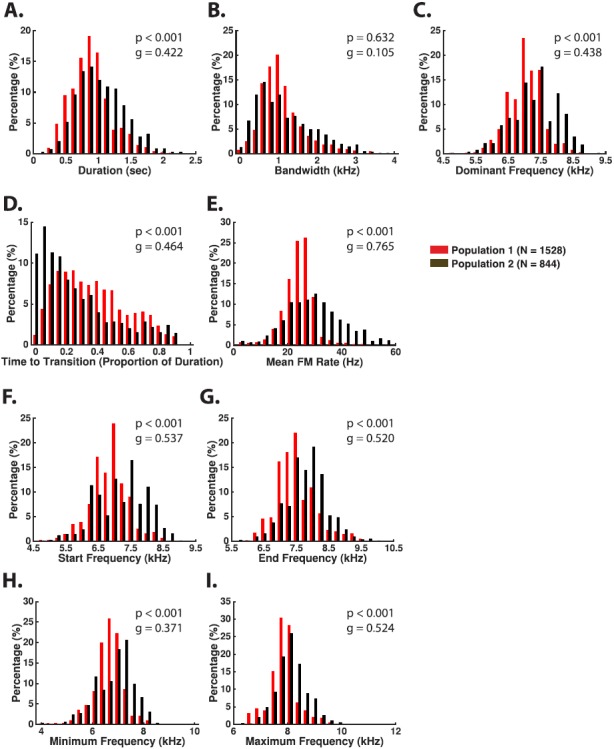
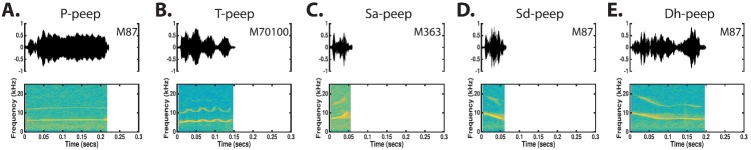
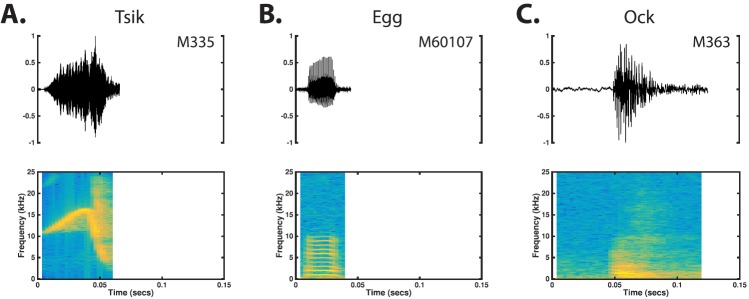
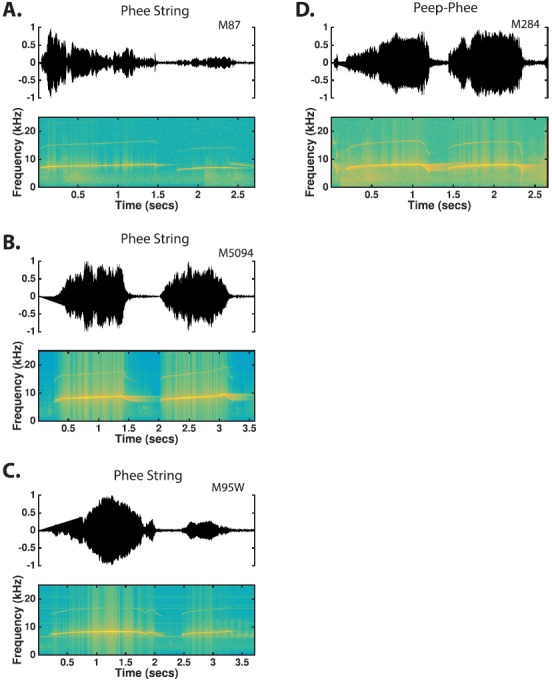
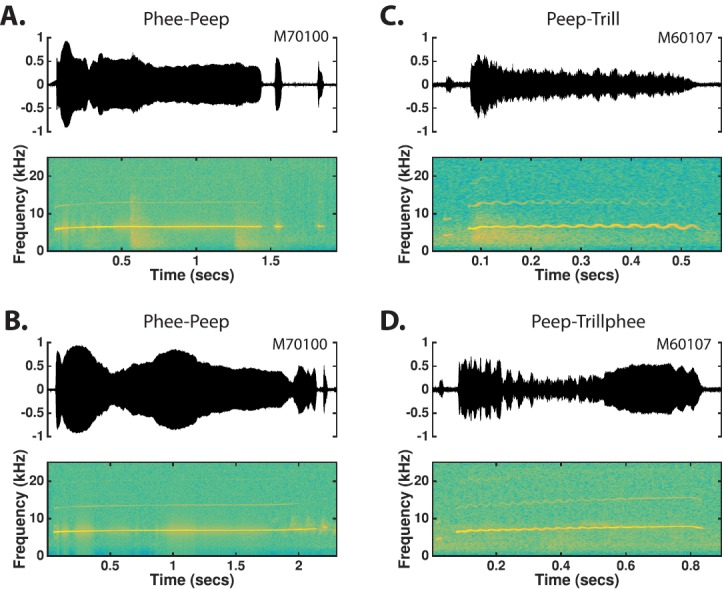
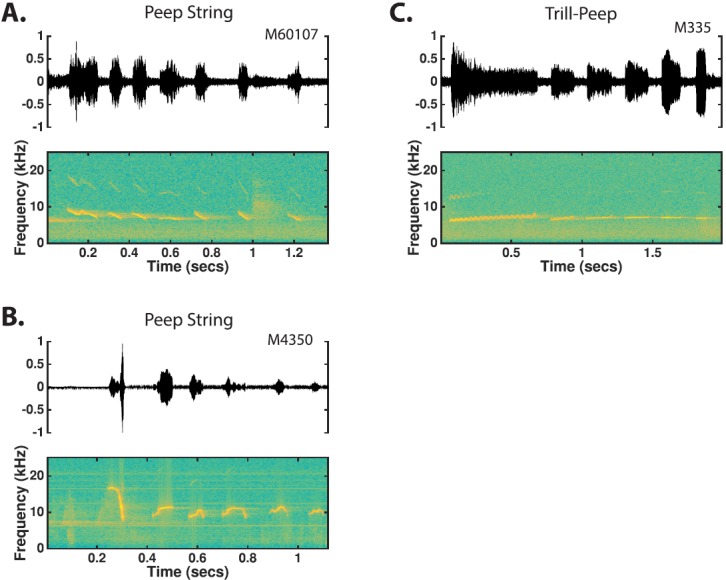
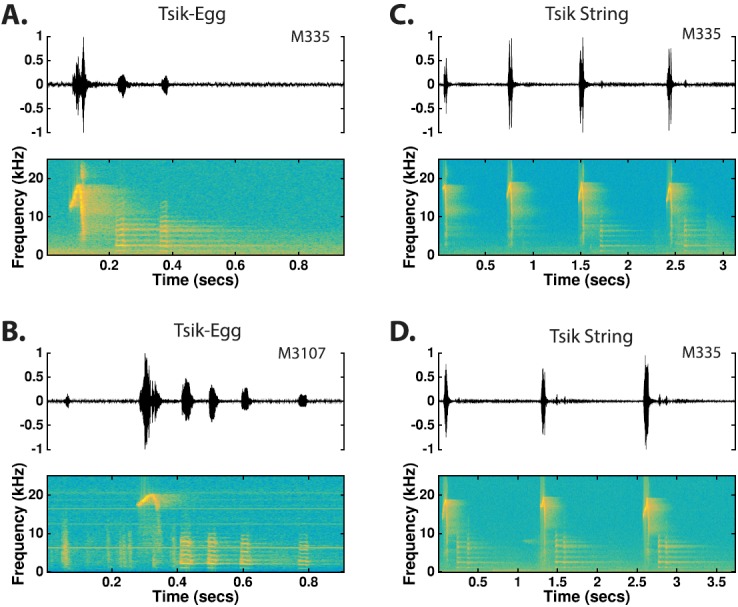
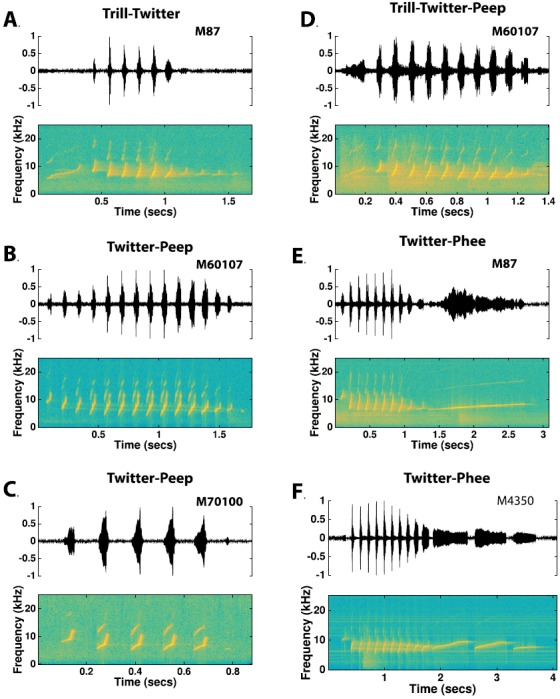
The common marmoset (Callithrix jacchus), a highly vocal New World primate species, has emerged in recent years as a promising animal model for studying brain mechanisms underlying perception, vocal production, and cognition. The present study provides a quantitative acoustic analysis of a large number of vocalizations produced by marmosets in a social environment within a captive colony. Previous classifications of the marmoset vocal repertoire were mostly based on qualitative observations. In the present study a variety of vocalizations from individually identified marmosets were sampled and multiple acoustic features of each type of vocalization were measured. Results show that marmosets have a complex vocal repertoire in captivity that consists of multiple vocalization types, including both simple calls and compound calls composed of sequences of simple calls. A detailed quantification of the vocal repertoire of the marmoset can serve as a solid basis for studying the behavioral significance of their vocalizations and is essential for carrying out studies that investigate such properties as perceptual boundaries between call types and among individual callers as well as neural coding mechanisms for vocalizations. It can also serve as the basis for evaluating abnormal vocal behaviors resulting from diseases or genetic manipulations.
Figures
References
-
- Bezerra, B. M. , and Souto, A. (2008). “ Structure and usage of the vocal repertoire of Calltihrix jacchus,” Int. J. Primatol. 29, 671–701.10.1007/s10764-008-9250-0 - DOI
-
- Bradbury, J. W. , and Vehrencamp, S. L. (1998). Principles of Animal Communication ( Sinauer Associates, Sunderland, MA: ), 882 pp.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous