

The Prodomain-bound Form of Bone Morphogenetic Protein 10 Is Biologically Active on Endothelial Cells
- PMID: 26631724
- PMCID: PMC4742757
- DOI: 10.1074/jbc.M115.683292
The Prodomain-bound Form of Bone Morphogenetic Protein 10 Is Biologically Active on Endothelial Cells
Abstract
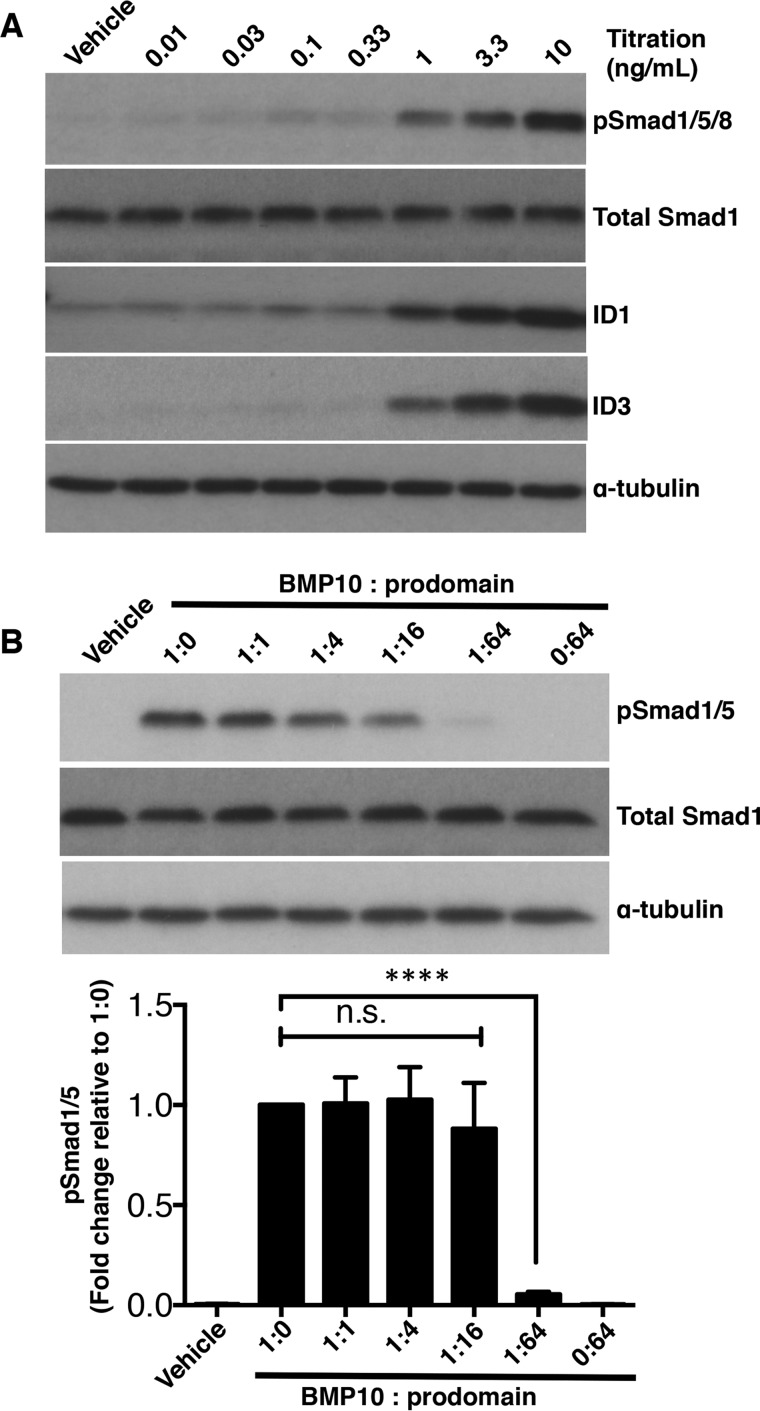
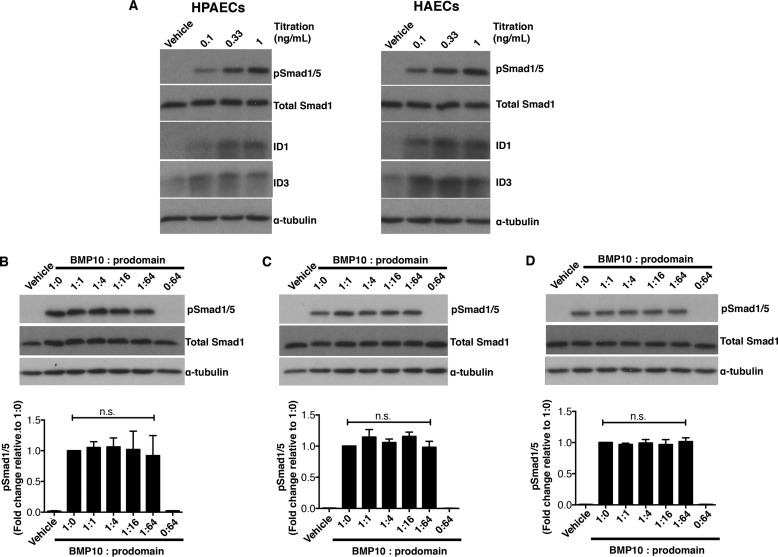
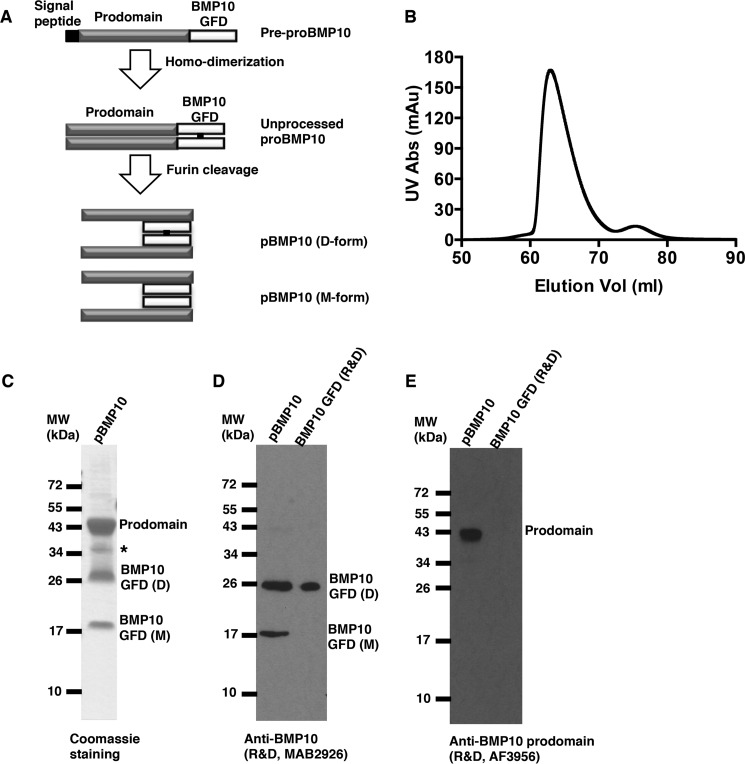
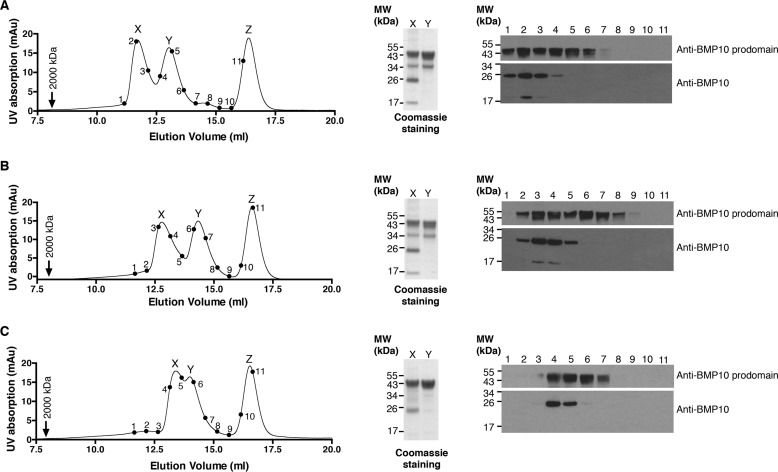
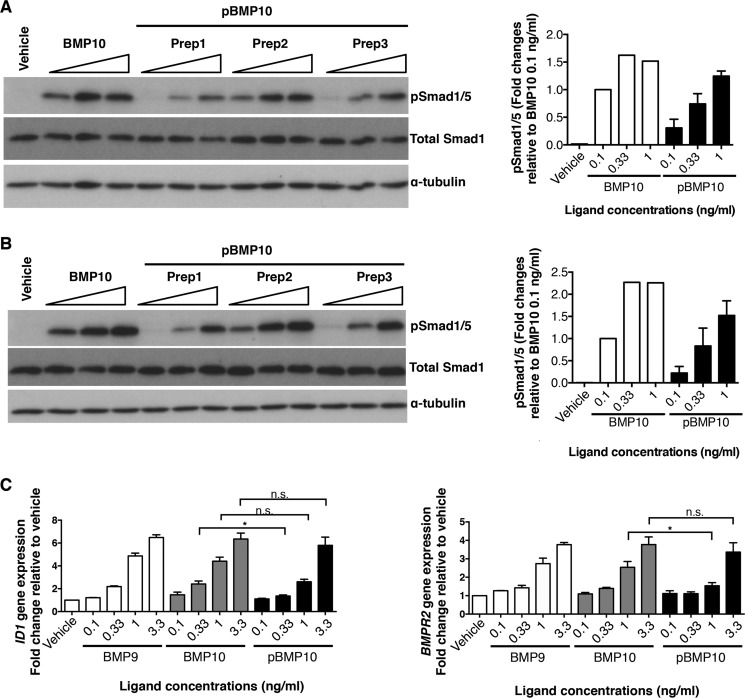
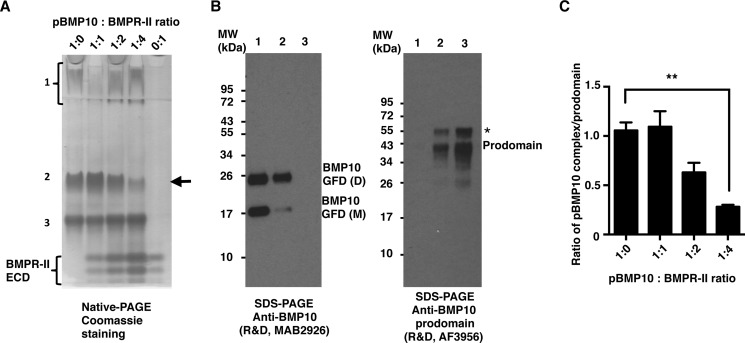
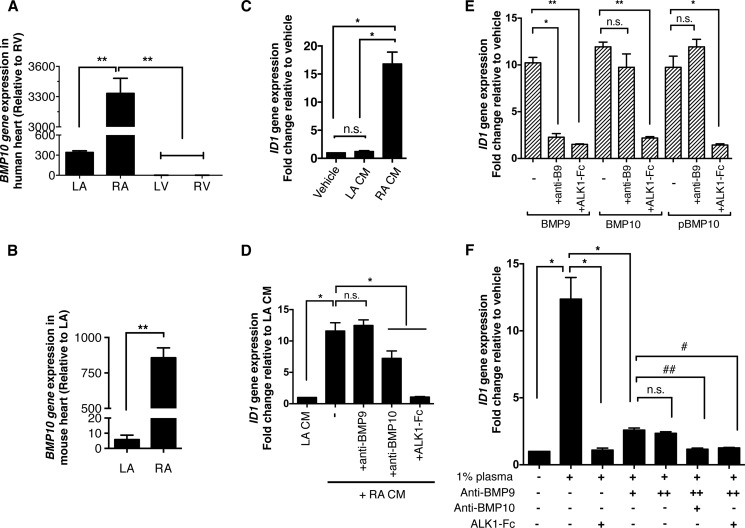
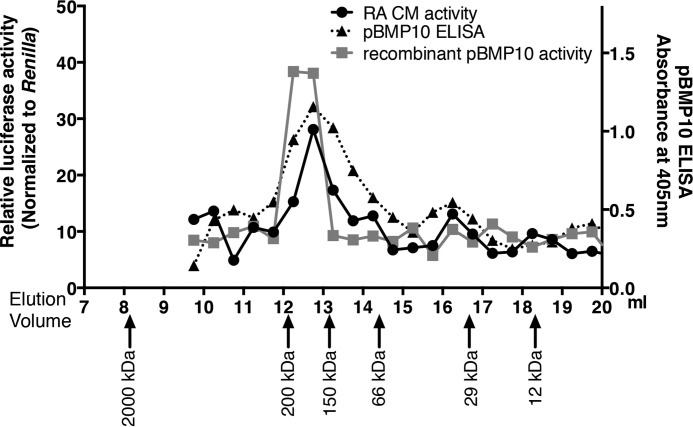
BMP10 is highly expressed in the developing heart and plays essential roles in cardiogenesis. BMP10 deletion in mice results in embryonic lethality because of impaired cardiac development. In adults, BMP10 expression is restricted to the right atrium, though ventricular hypertrophy is accompanied by increased BMP10 expression in a rat hypertension model. However, reports of BMP10 activity in the circulation are inconclusive. In particular, it is not known whether in vivo secreted BMP10 is active or whether additional factors are required to achieve its bioactivity. It has been shown that high-affinity binding of the BMP10 prodomain to the mature ligand inhibits BMP10 signaling activity in C2C12 cells, and it was proposed that prodomain-bound BMP10 (pBMP10) complex is latent. In this study, we demonstrated that the BMP10 prodomain did not inhibit BMP10 signaling activity in multiple endothelial cells, and that recombinant human pBMP10 complex, expressed in mammalian cells and purified under native conditions, was fully active. In addition, both BMP10 in human plasma and BMP10 secreted from the mouse right atrium were fully active. Finally, we confirmed that active BMP10 secreted from mouse right atrium was in the prodomain-bound form. Our data suggest that circulating BMP10 in adults is fully active and that the reported vascular quiescence function of BMP10 in vivo is due to the direct activity of pBMP10 and does not require an additional activation step. Moreover, being an active ligand, recombinant pBMP10 may have therapeutic potential as an endothelial-selective BMP ligand, in conditions characterized by loss of BMP9/10 signaling.
Keywords: bone morphogenetic protein (BMP); bone morphogenetic protein 10 (BMP10); cell biology; endothelial cell; signal transduction; transforming growth factor beta (TGF-β).
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Neuhaus H., Rosen V., and Thies R. S. (1999) Heart specific expression of mouse BMP-10 a novel member of the TGF-β superfamily. Mech. Dev. 80, 181–184 - PubMed
-
- Chen H., Yong W., Ren S., Shen W., He Y., Cox K. A., Zhu W., Li W., Soonpaa M., Payne R. M., Franco D., Field L. J., Rosen V., Wang Y., and Shou W. (2006) Overexpression of bone morphogenetic protein 10 in myocardium disrupts cardiac postnatal hypertrophic growth. J. Biol. Chem. 281, 27481–27491 - PMC - PubMed
-
- Nakano N., Hori H., Abe M., Shibata H., Arimura T., Sasaoka T., Sawabe M., Chida K., Arai T., Nakahara K., Kubo T., Sugimoto K., Katsuya T., Ogihara T., Doi Y., Izumi T., and Kimura A. (2007) Interaction of BMP10 with Tcap may modulate the course of hypertensive cardiac hypertrophy. Am. J. Physiol. Heart Circ. Physiol. 293, H3396–H3403 - PubMed
-
- Mazerbourg S., Sangkuhl K., Luo C. W., Sudo S., Klein C., and Hsueh A. J. (2005) Identification of receptors and signaling pathways for orphan bone morphogenetic protein/growth differentiation factor ligands based on genomic analyses. J. Biol. Chem. 280, 32122–32132 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
