

Minor Loops in Major Folds: Enhancer-Promoter Looping, Chromatin Restructuring, and Their Association with Transcriptional Regulation and Disease
- PMID: 26632825
- PMCID: PMC4669122
- DOI: 10.1371/journal.pgen.1005640
Minor Loops in Major Folds: Enhancer-Promoter Looping, Chromatin Restructuring, and Their Association with Transcriptional Regulation and Disease
Abstract
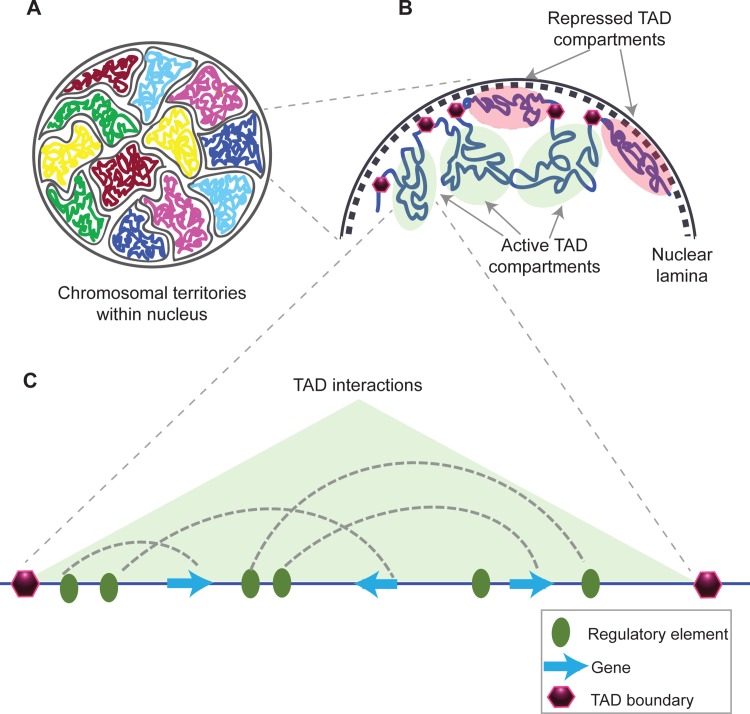
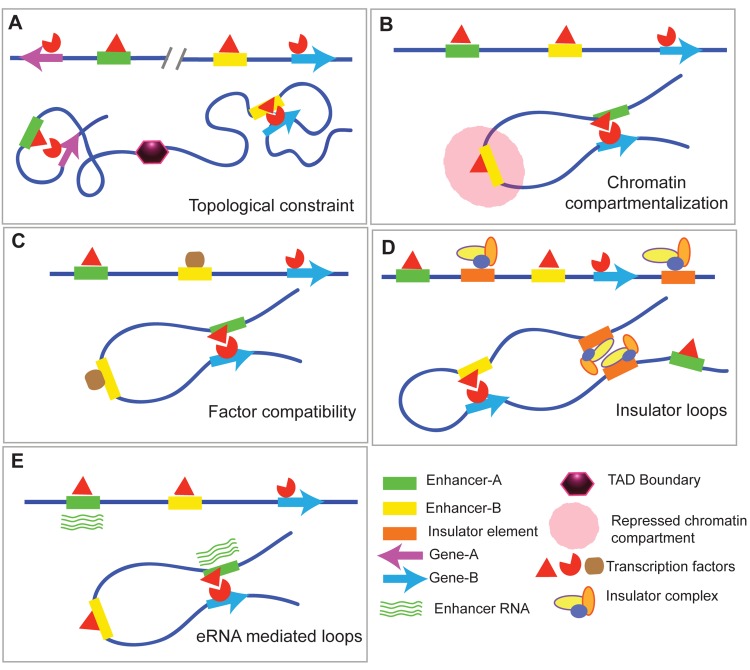
The organization and folding of chromatin within the nucleus can determine the outcome of gene expression. Recent technological advancements have enabled us to study chromatin interactions in a genome-wide manner at high resolution. These studies have increased our understanding of the hierarchy and dynamics of chromatin domains that facilitate cognate enhancer-promoter looping, defining the transcriptional program of different cell types. In this review, we focus on vertebrate chromatin long-range interactions as they relate to transcriptional regulation. In addition, we describe how the alteration of boundaries that mark discrete regions in the genome with high interaction frequencies within them, called topological associated domains (TADs), could lead to various phenotypes, including human diseases, which we term as "TADopathies."
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
- 1R01NS079231/NS/NINDS NIH HHS/United States
- R01 CA197139/CA/NCI NIH HHS/United States
- GM61390/GM/NIGMS NIH HHS/United States
- P30 DK098722/DK/NIDDK NIH HHS/United States
- U01 GM061390/GM/NIGMS NIH HHS/United States
- U19 GM061390/GM/NIGMS NIH HHS/United States
- R01 DK090382/DK/NIDDK NIH HHS/United States
- 1R01HG006768/HG/NHGRI NIH HHS/United States
- 1R01DK090382/DK/NIDDK NIH HHS/United States
- R01 NS079231/NS/NINDS NIH HHS/United States
- 1R01CA197139/CA/NCI NIH HHS/United States
- R01 HG006768/HG/NHGRI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
