

The most sensitive inputs to cutaneous representing regions of primary somatosensory cortex do not change with behavioral training
- PMID: 26634900
- PMCID: PMC4760438
- DOI: 10.14814/phy2.12623
The most sensitive inputs to cutaneous representing regions of primary somatosensory cortex do not change with behavioral training
Abstract
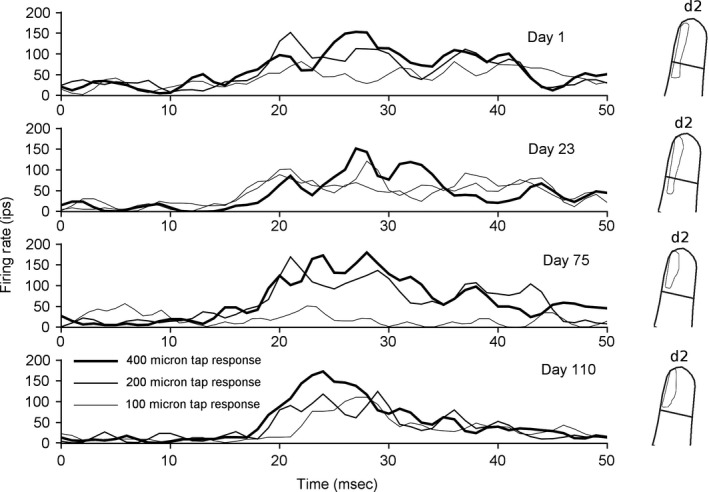
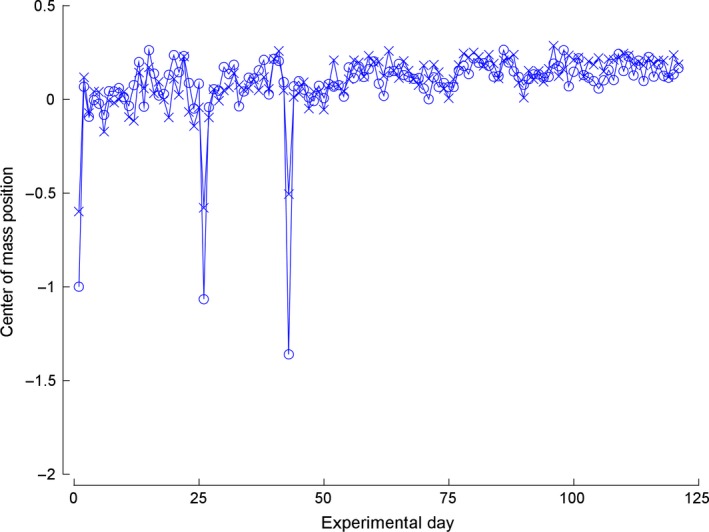
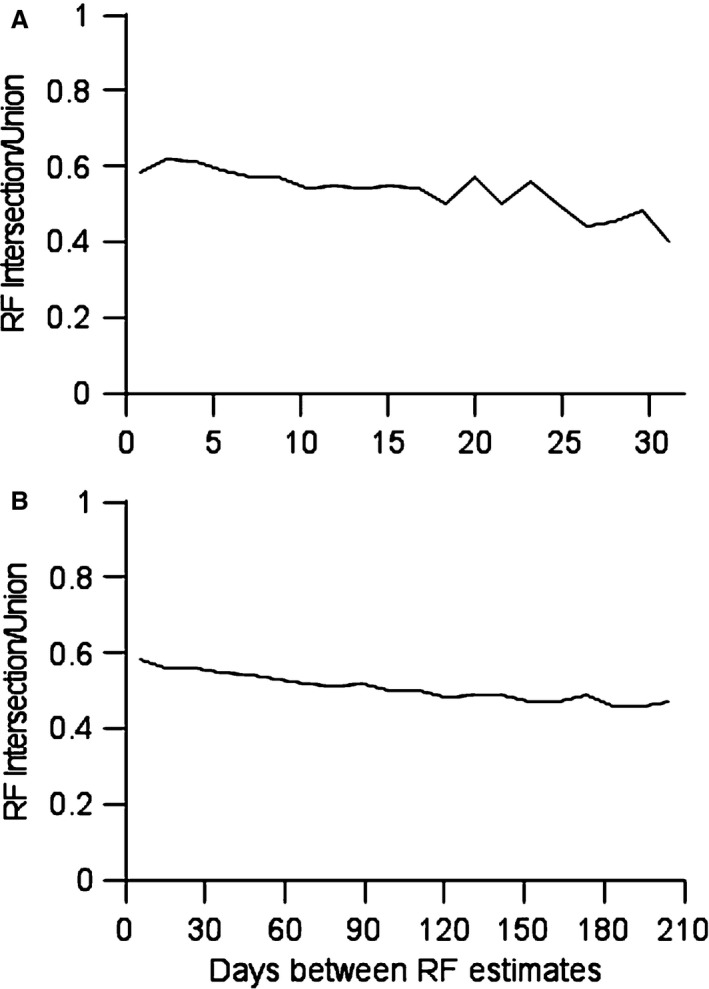
Learning a sensory detection task leads to an increased primary sensory cortex response to the detected stimulus, while learning a sensory discrimination task additionally leads to a decreased sensory cortex response to the distractor stimulus. Neural responses are scaled up, and down, in strength, along with concomitant changes in receptive field size. The present work considers neural response properties that are invariant to learning. Data are drawn from two animals that were trained to detect and discriminate spatially separate taps delivered to positions on the skin of their fingers. Each animal was implanted with electrodes positioned in area 3b, and responses were derived on a near daily basis over 84 days in animal 1 and 202 days in animal 2. Responses to taps delivered in the receptive field were quantitatively measured each day, and receptive fields were audiomanually mapped each day. In the subset of responses that had light cutaneous receptive fields, a preponderance of the days, the most sensitive region of the field was invariant to training. This skin region was present in the receptive field on all, or nearly all, occasions in which the receptive field was mapped, and this region constituted roughly half of the most sensitive region. These results suggest that maintaining the most sensitive inputs as dominant in cortical receptive fields provide a measure of stability that may be transformationally useful for minimizing reconstruction errors and perceptual constancy.
Keywords: Implant; map plasticity; receptive field; somatosensory.
© 2015 The Authors. Physiological Reports published by Wiley Periodicals, Inc. on behalf of the American Physiological Society and The Physiological Society.
Figures

Similar articles
-
Topographic reorganization of the hand representation in cortical area 3b owl monkeys trained in a frequency-discrimination task.J Neurophysiol. 1992 May;67(5):1031-56. doi: 10.1152/jn.1992.67.5.1031. J Neurophysiol. 1992. PMID: 1597696
-
Frequency discrimination training engaging a restricted skin surface results in an emergence of a cutaneous response zone in cortical area 3a.J Neurophysiol. 1992 May;67(5):1057-70. doi: 10.1152/jn.1992.67.5.1057. J Neurophysiol. 1992. PMID: 1597697
-
Experience-dependent plasticity in S1 caused by noncoincident inputs.J Neurophysiol. 2005 Sep;94(3):2239-50. doi: 10.1152/jn.00172.2005. J Neurophysiol. 2005. PMID: 16105958 Free PMC article.
-
Intracortical processes regulating the integration of sensory information.Prog Brain Res. 1990;86:129-41. doi: 10.1016/s0079-6123(08)63172-6. Prog Brain Res. 1990. PMID: 1982365 Review.
-
Physiological memory in primary auditory cortex: characteristics and mechanisms.Neurobiol Learn Mem. 1998 Jul-Sep;70(1-2):226-51. doi: 10.1006/nlme.1998.3850. Neurobiol Learn Mem. 1998. PMID: 9753599 Review.
References
-
- Allard, T. , Clark S. A., Jenkins W. M., and Merzenich M. M.. 1991. Reorganization of somatosensory area 3b representations in adult owl monkeys after digital syndactyly. J. Neurophysiol. 66:1048–1058. - PubMed
-
- Basbaum, A. I. , and Wall P. D.. 1976. Chronic changes in the response of cells in adult cat dorsal horn following partial deafferentation: the appearance of responding cells in a previously non‐responsive region. Brain Res. 116:181–204. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
