

Drosophila Bitter Taste(s)
- PMID: 26635553
- PMCID: PMC4658422
- DOI: 10.3389/fnint.2015.00058
Drosophila Bitter Taste(s)
Abstract
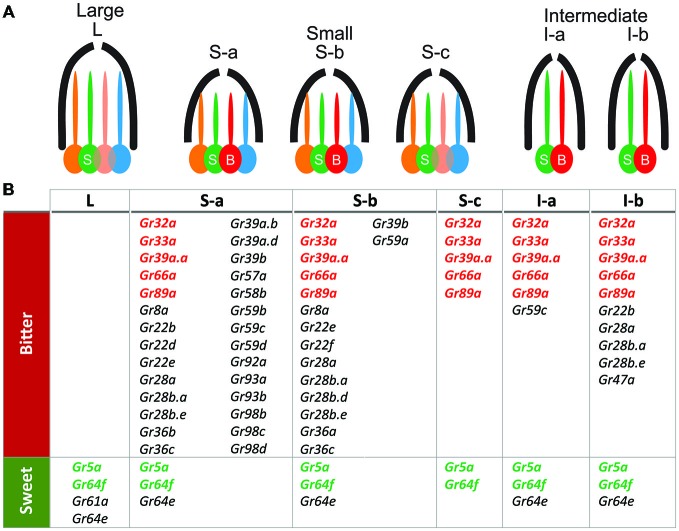
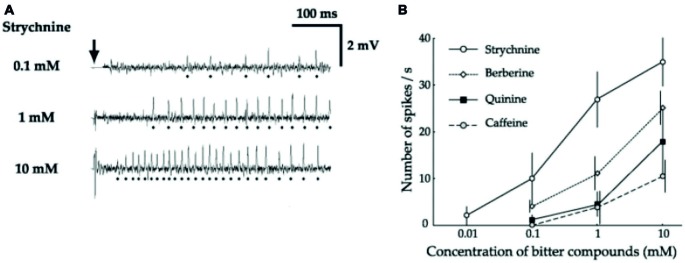
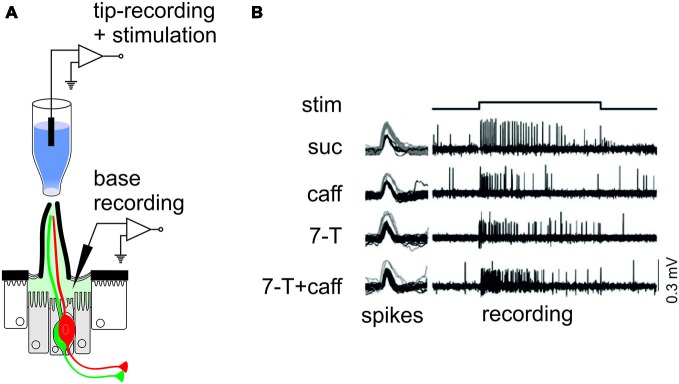
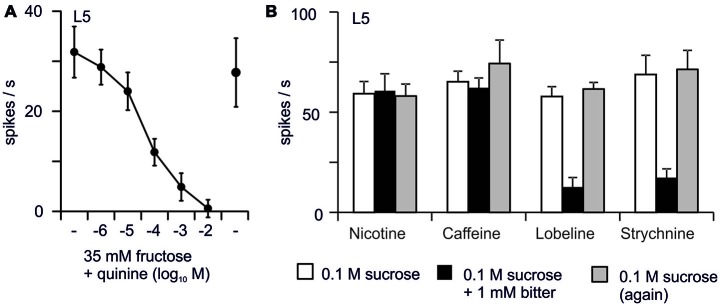
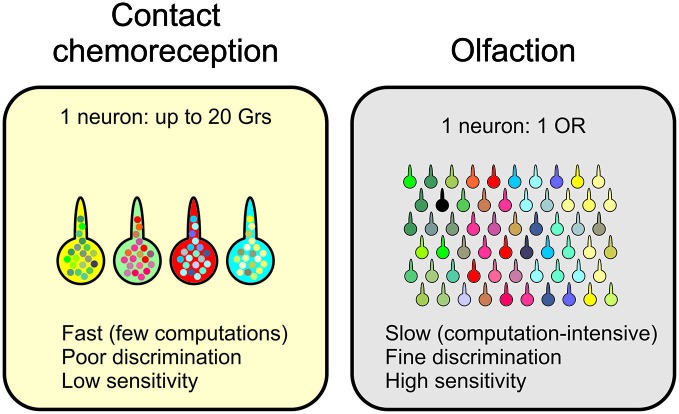
Most animals possess taste receptors neurons detecting potentially noxious compounds. In humans, the ligands which activate these neurons define a sensory space called "bitter". By extension, this term has been used in animals and insects to define molecules which induce aversive responses. In this review, based on our observations carried out in Drosophila, we examine how bitter compounds are detected and if bitter-sensitive neurons respond only to molecules bitter to humans. Like most animals, flies detect bitter chemicals through a specific population of taste neurons, distinct from those responding to sugars or to other modalities. Activating bitter-sensitive taste neurons induces aversive reactions and inhibits feeding. Bitter molecules also contribute to the suppression of sugar-neuron responses and can lead to a complete inhibition of the responses to sugar at the periphery. Since some bitter molecules activate bitter-sensitive neurons and some inhibit sugar detection, bitter molecules are represented by two sensory spaces which are only partially congruent. In addition to molecules which impact feeding, we recently discovered that the activation of bitter-sensitive neurons also induces grooming. Bitter-sensitive neurons of the wings and of the legs can sense chemicals from the gram negative bacteria, Escherichia coli, thus adding another biological function to these receptors. Bitter-sensitive neurons of the proboscis also respond to the inhibitory pheromone, 7-tricosene. Activating these neurons by bitter molecules in the context of sexual encounter inhibits courting and sexual reproduction, while activating these neurons with 7-tricosene in a feeding context will inhibit feeding. The picture that emerges from these observations is that the taste system is composed of detectors which monitor different "categories" of ligands, which facilitate or inhibit behaviors depending on the context (feeding, sexual reproduction, hygienic behavior), thus considerably extending the initial definition of "bitter" tasting.
Keywords: aversive; behavior; electrophysiology; insects; pheromones; taste.
Figures
References
-
- Altner H., Prillinger L. (1980). Ultrastructure of invertebrate chemo-, thermo- and hygroreceptors and its functional significance. Int. Rev. Cytol. 67, 69–139. 10.1016/s0074-7696(08)62427-4 - DOI
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases