

Automatic Sleep Spindle Detection and Genetic Influence Estimation Using Continuous Wavelet Transform
- PMID: 26635577
- PMCID: PMC4652604
- DOI: 10.3389/fnhum.2015.00624
Automatic Sleep Spindle Detection and Genetic Influence Estimation Using Continuous Wavelet Transform
Abstract
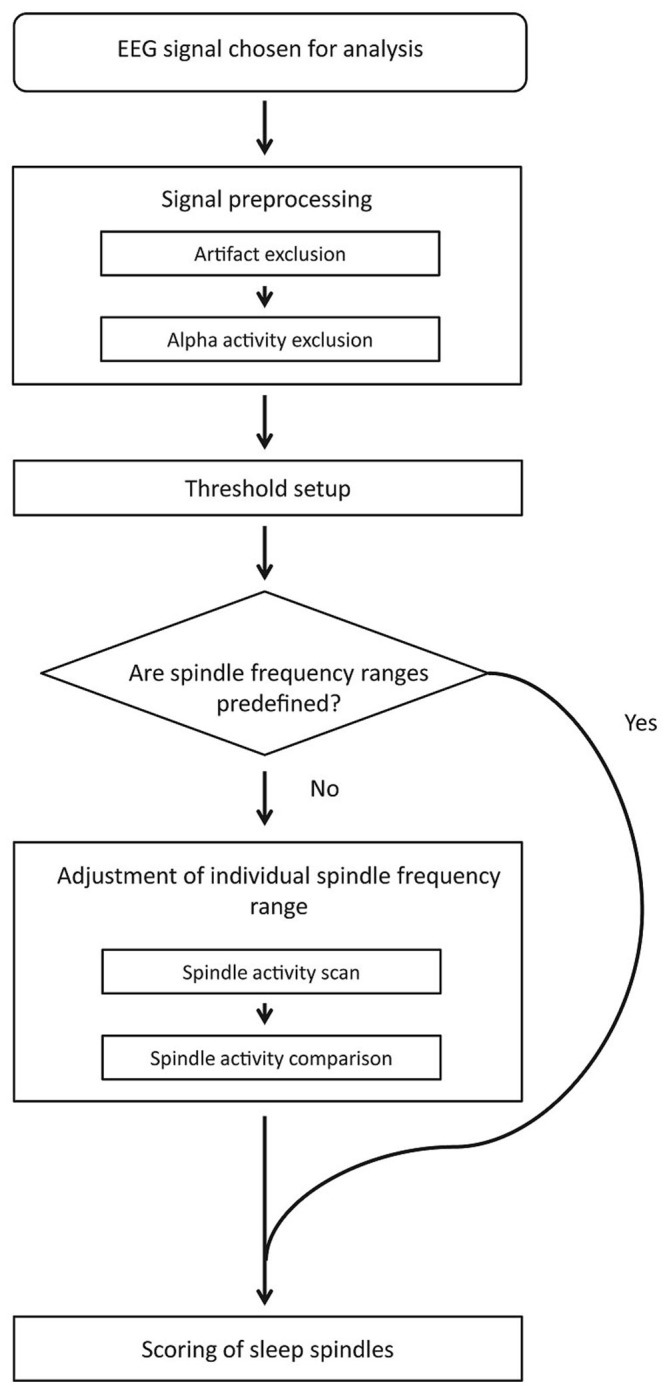
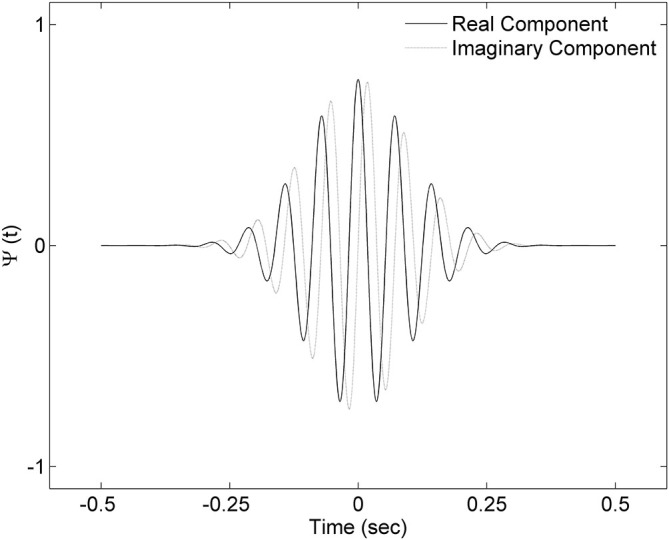
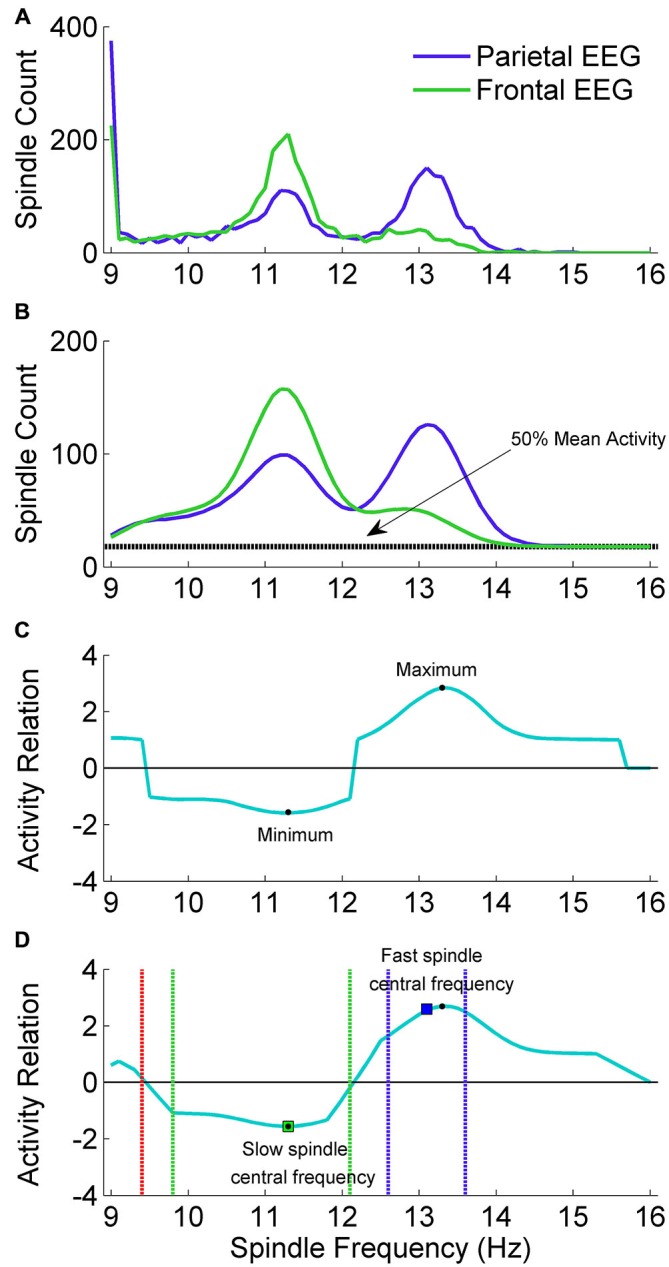
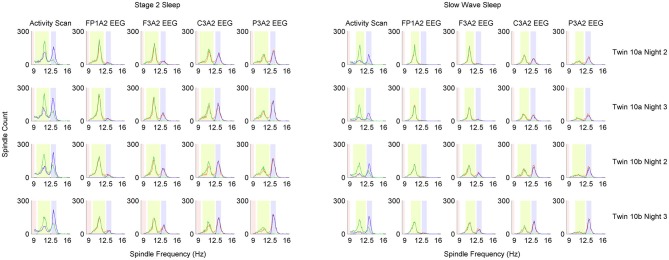
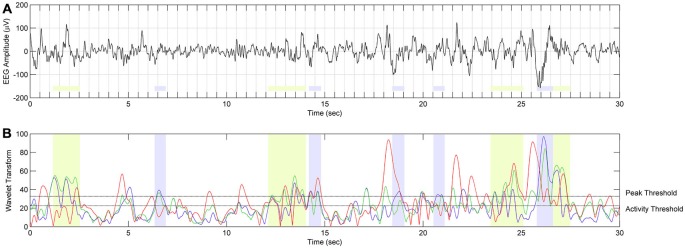
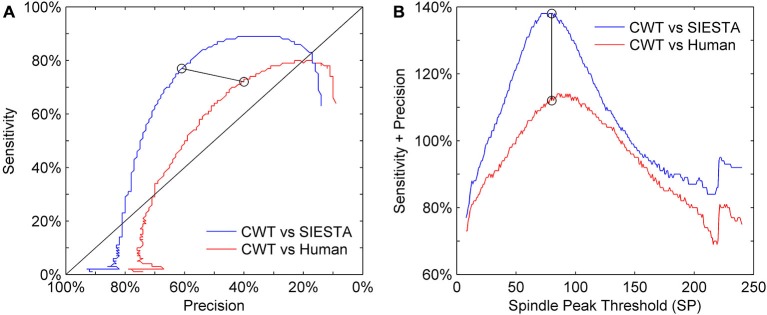
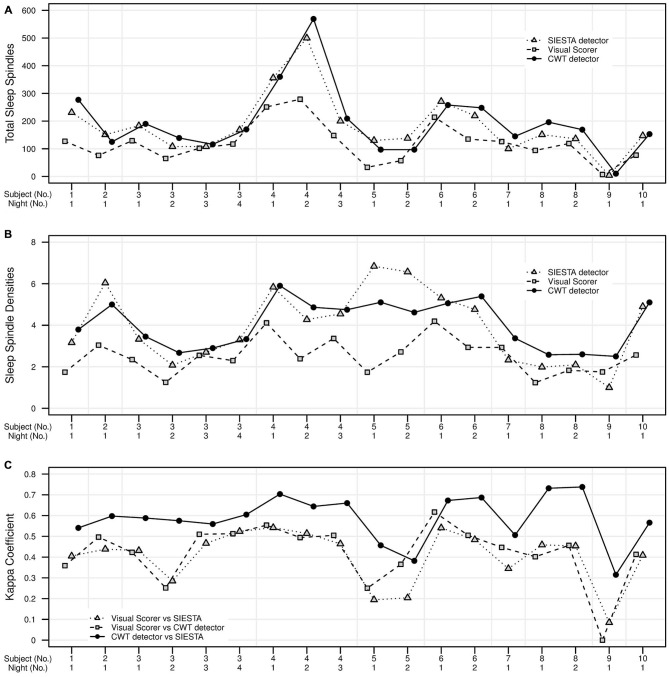
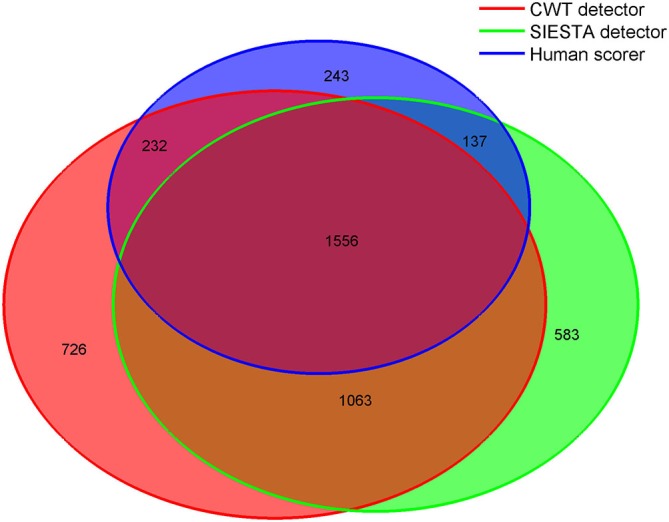
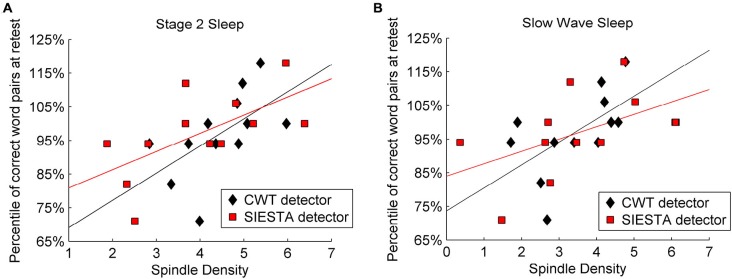
Mounting evidence for the role of sleep spindles in neuroplasticity has led to an increased interest in these non-rapid eye movement (NREM) sleep oscillations. It has been hypothesized that fast and slow spindles might play a different role in memory processing. Here, we present a new sleep spindle detection algorithm utilizing a continuous wavelet transform (CWT) and individual adjustment of slow and fast spindle frequency ranges. Eighteen nap recordings of ten subjects were used for algorithm validation. Our method was compared with both a human scorer and a commercially available SIESTA spindle detector. For the validation set, mean agreement between our detector and human scorer measured during sleep stage 2 using kappa coefficient was 0.45, whereas mean agreement between our detector and SIESTA algorithm was 0.62. Our algorithm was also applied to sleep-related memory consolidation data previously analyzed with a SIESTA detector and confirmed previous findings of significant correlation between spindle density and declarative memory consolidation. We then applied our method to a study in monozygotic (MZ) and dizygotic (DZ) twins, examining the genetic component of slow and fast sleep spindle parameters. Our analysis revealed strong genetic influence on variance of all slow spindle parameters, weaker genetic effect on fast spindles, and no effects on fast spindle density and number during stage 2 sleep.
Keywords: EEG; automatic detection; heritability; sleep spindle; twins.
Figures
References
-
- Addison P. (2002). The Illustrated Wavelet Transform Handbook: Introductory Theory and Applications in Science, Engineering, Medicine and Finance, Bristol, Philadelphia: Institute of Physics Publishing.
-
- Anderer P., Gruber G., Parapatics S., Woertz M., Miazhynskaia T., Klosch G., et al. (2005). An E-health solution for automatic sleep classification according to Rechtschaffen and Kales: validation study of the Somnolyzer 24 × 7 utilizing the Siesta database. Neuropsychobiology 51, 115–133. 10.1159/000085205 - DOI - PubMed
-
- Anderer P., Klösch G., Gruber G., Trenker E., Pascual-Marqui R. D., Zeitlhofer J., et al. (2001). Low-resolution brain electromagnetic tomography revealed simultaneously active frontal and parietal sleep spindle sources in the human cortex. Neuroscience 103, 581–592. 10.1016/s0306-4522(01)00028-8 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials