

Maturation and Functional Integration of New Granule Cells into the Adult Hippocampus
- PMID: 26637288
- PMCID: PMC4691791
- DOI: 10.1101/cshperspect.a018903
Maturation and Functional Integration of New Granule Cells into the Adult Hippocampus
Abstract
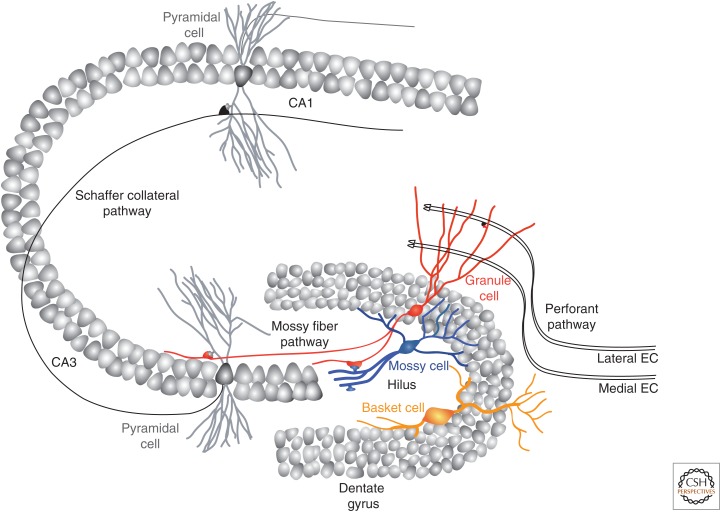
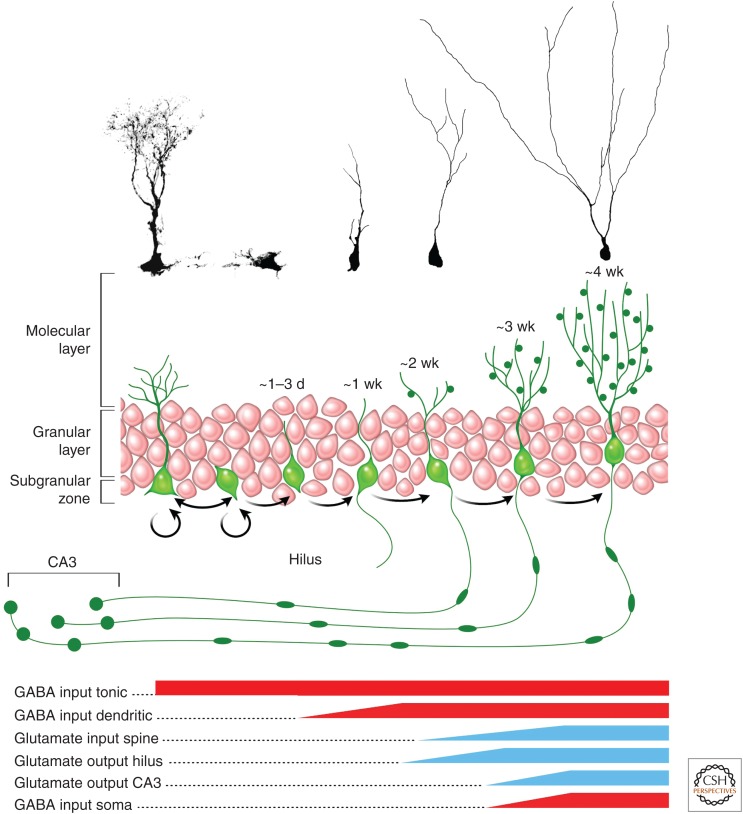
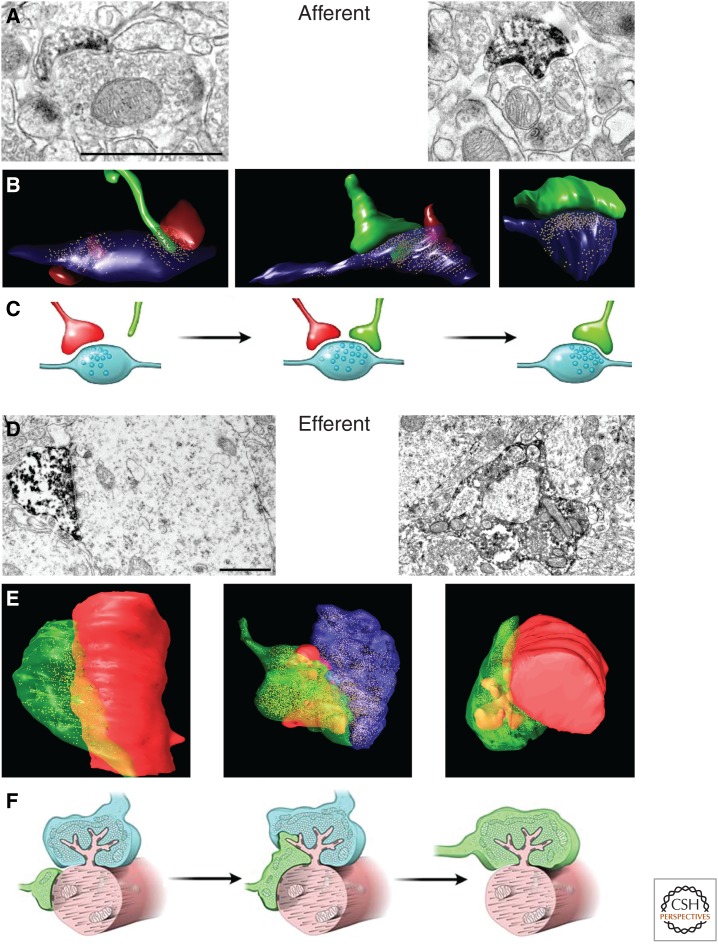
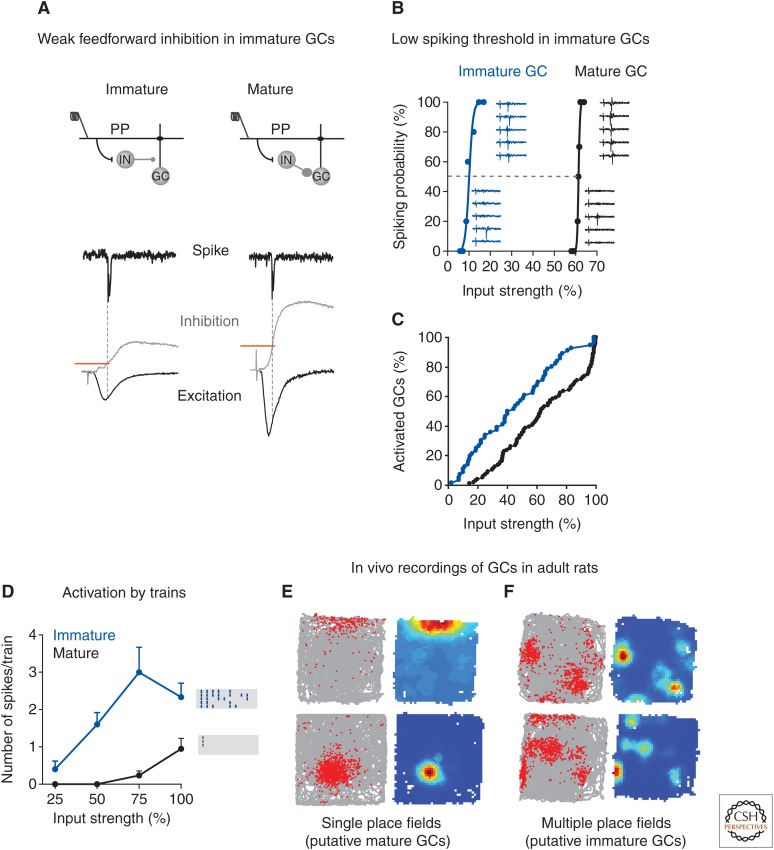
The adult hippocampus generates functional dentate granule cells (GCs) that release glutamate onto target cells in the hilus and cornus ammonis (CA)3 region, and receive glutamatergic and γ-aminobutyric acid (GABA)ergic inputs that tightly control their spiking activity. The slow and sequential development of their excitatory and inhibitory inputs makes them particularly relevant for information processing. Although they are still immature, new neurons are recruited by afferent activity and display increased excitability, enhanced activity-dependent plasticity of their input and output connections, and a high rate of synaptogenesis. Once fully mature, new GCs show all the hallmarks of neurons generated during development. In this review, we focus on how developing neurons remodel the adult dentate gyrus and discuss key aspects that illustrate the potential of neurogenesis as a mechanism for circuit plasticity and function.
Copyright © 2016 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Amaral DG. 1978. A Golgi study of cell types in the hilar region of the hippocampus in the rat. J Comp Neurol 182: 851–914. - PubMed
-
- Amaral DG, Ishizuka N, Claiborne B. 1990. Neurons, numbers and the hippocampal network. Prog Brain Res 83: 1–11. - PubMed
-
- Barria A, Malinow R. 2005. NMDA receptor subunit composition controls synaptic plasticity by regulating binding to CaMKII. Neuron 48: 289–301. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous